Dissociable Neural Mechanisms for Encoding
Predictable and Unpredictable Events
Christopher Summerfield and Jennifer A. Mangels
Abstracto
& Attention is a necessary condition for the formation of new
episodic memories, yet little is known about how dissociable
attentional mechanisms for ‘‘top-down’’ and ‘‘bottom-up’’
orienting contribute to encoding. Aquí, subjects performed an
intentional encoding task in which to-be-learned items were
interspersed with irrelevant stimuli such that subjects could
anticipate the appearance of some study items but not others.
Subjects were more likely to later remember stimuli whose
appearance was predictable at encoding. Electroencephalo-
graphic data were acquired during the study phase of the
experiment to assess how synchronous neural activity related
to later memory for predictable stimuli (to which attention
could be oriented in a top-down fashion) and unpredictable
estímulos (which rely to a greater extent on bottom-up atten-
tional orienting). Over left frontal regions, gamma-band activity
(25–55 Hz) early ((cid:1)150 mseg) in the epoch was a robust pre-
dictor of later memory for predictable items, consistent with an
emerging view that links high-frequency neural synchrony to
top-down attention. Por el contrario, más tarde ((cid:1)400 mseg) theta-band
actividad (4–8 Hz) over the left and midline frontal cortex pre-
dicted subsequent memory for unpredictable items, sugerencia
a role in bottom-up attentional orienting. These results reveal
for the first time the contribution of dissociable attentional
mechanisms to successful encoding and contribute to a grow-
ing literature dedicated to understanding the role of neural
synchrony in cognition. &
INTRODUCCIÓN
Considerable advances in our understanding of the neu-
ral correlates of episodic memory formation are owed
to an experimental paradigm known as the ‘‘difference
of memory’’ (DM) paradigma (Paller, Kutas, & Mayes,
1987; Sanquist, Rohrbaugh, Syndulko, & Lindsley, 1980).
In combination with event-related neuroimaging tech-
niques, the DM paradigm allows the back-sorting of
study-phase neural activity on the basis of performance
on a subsequent retrieval test. Items that were pre-
sented at study thus can be classified into those that
were successfully encoded and those that were not, y
patterns of brain activity associated with these two types
of trial can then be compared. Using event-related func-
tional magnetic resonance imaging (resonancia magnética funcional), the DM
paradigm has been used both to confirm the involve-
ment of medial temporal lobe structures identified by
early lesion work (Scoville & Milner, 1957), and to de-
fine neocortical regions, notably those in the inferior
frontal and parietal lobes, which contribute to success-
ful encoding (Rugg, Otten, & Henson, 2002; Buckner,
logan, Donaldson, & Rodador, 2000; Wagner et al.,
1998). The DM paradigm has also been used with
event-related potentials (ERPs) to reveal an unfold-
ing sequence of encoding-related components locked
Columbia University
to the onset of the stimulus: Por ejemplo, an early neg-
ativity over left frontotemporal sites (Mangels, Picton,
& Craik, 2001), a positive-going parietal component
at 300–600 msec (Otten & Donchin, 2000; Paller et al.,
1987), and a late sustained positivity at frontal elec-
trodes (Fabiani, Karis, & Donchin, 1990). Similarmente, suc-
cessful encoding has been associated with increased
electroencephalographic (EEG) synchrony in the theta
(4–8 Hz) (Summerfield & Mangels, 2005a; Montaña de cedro,
Kahana, Howard, Donner, & Madsen, 2003; Klimesch,
Doppelmayr, Russegger, & Pachinger, 1996) and gam-
mamá (>25 Hz) bands, particularly over anterior neocor-
tical sites (Gruber, Tsivilis, Montaldi, & Muller, 2004;
Sederberg et al., 2003) and within the medial temporal
lobe (Fell, Klaver, Elfadil, et al., 2003; Fell, Klaver,
Lehnertz, et al., 2001).
The DM paradigm capitalizes on moment-by-moment
variability in the likelihood that our perceptual experi-
ences will be laid down as new traces in long-term
memory: that although some items will be successfully
encoded, others will not. Crucial to our understanding
of how new memories are formed, de este modo, is an under-
standing of the sources of variability in encoding suc-
impuesto. Factors intrinsic to the events we experience are an
important predictor of
later memory. Por ejemplo,
stimuli that are novel or distinctive by virtue of their
perceptual, conceptual, or emotional attributes enjoy
a well-described advantage during learning (the Von
D 2006 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 18:7, páginas. 1120–1132
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Restorff effect) (Strange, Otten, Josephs, Rugg, & Dolan,
2002; Fabiani & Donchin, 1995; von Restorff, 1933).
Sin embargo, extrinsic factors, such as the depth of concep-
tual processing to which the study material is subjected
(Craik & Lockhart, 1972) or the level of attentional
resources available during encoding (Craik, Govoni,
Naveh-Benjamin, & anderson, 1996) also have a power-
ful modulatory role on both memory performance and
neural activity that accompanies successful
aprendiendo
(Kensinger, Clarke, & Corkin, 2003; Mangels et al.,
2001; Fletcher, Shallice, & Dolan, 1998).
The relationship between attention and encoding is of
particular interest to researchers studying the neurobi-
ology of memory, as it has been shown that unattended
items typically cannot be later recalled or consciously
recollected (Craik et al., 1996; Gardiner & Parkin, 1990).
Given that attention is a necessary condition for epi-
sodic encoding, it is likely that a subset of the brain
activity found to predict later memory in the DM para-
digm reflects the orienting of attention to the study
material, independent of other neural phenomena that
may vary with stimulus distinctiveness,
level of con-
ceptual processing, or the formation of the declarative
memory trace itself. Respectivamente, some previous DM
studies have divided attention during the study phase
in an attempt to identify neural correlates of successful
encoding that exhibit a parallel sensitivity to reduction in
attentional load (Kensinger et al., 2003; Mangels et al.,
2001; Anderson et al., 2000; Iidaka, anderson, Kapur,
Cabeza, & Craik, 2000). One result that has consistently
emerged from these studies is that activity in the left
inferior prefrontal cortex, which is perhaps the neocor-
tical zone most consistently implicated in successful en-
coding under full attention conditions, does not predict
later memory when an attention-demanding secondary
task is imposed at study. One corollary of this finding is
that reductions in attentional resources impair later
memory at least in part because under divided attention,
left inferior frontal regions are no longer able to con-
tribute to encoding. Drawing upon these data, también
as upon evidence that left frontal cortical sites observed
to be active in DM studies overlap with those thought
to subserve semantic retrieval (Thompson-Schill, 2003)
and phonological rehearsal (Davachi, Maril, & Wagner,
2001), it has been proposed that dividing attention im-
pairs memory because it precludes elaborative concep-
tual processing (such as thinking about the meaning
of an item or rehearsing it in working memory) that rely
on these left anterior structures (Naveh-Benjamin, Guez,
& Marom, 2003; Naveh-Benjamin, Craik, Gavrilescu, &
anderson, 2000; Craik et al., 1996; Craik & Lockhart,
1972). This theory is consistent with the view that the
(attention dependent) function of the left prefrontal
cortex in successful encoding is to rehearse, associate,
or otherwise conceptually elaborate information in the
service of optimal encoding (Fletcher, Shallice, & Dolan,
2000).
Dividing attention at study offers insight into how en-
coding success varies under different levels of global pro-
cessing resources but is less well suited to exploring how
dissociable attentional networks for orienting and selec-
tion are involved in memory formation. Contemporary
theories describe attention as a modular system (Corbetta
& Shulman, 2002; Mesulam, 1998; posner & Petersen,
1990), with dissociable mechanisms for ‘‘top-down’’ (o
‘‘endogenous’’) and ‘‘bottom-up’’ (or ‘‘exogenous’’) ori-
enting. Top-down attention involves the selection of a
relevant feature, representación, or location from an
expected or predictable stimulus, whereas bottom-up
orienting occurs to stimuli that capture attention by
‘‘popping out’’ unexpectedly from an undifferentiated
contexto. It makes intuitive sense that these attentional
mechanisms may make different contributions to encod-
En g. Por ejemplo, during serial list learning, where items
follow each other in predictable succession, atención
can be reliably oriented in a top-down fashion to each
study item. Sin embargo, when an item occurs unexpectedly,
the degree to which attention can be disengaged from
the current task and reoriented to the novel stimulus in
a controlled fashion is likely to be a good predictor of
aprendiendo.
The object of this study was to compare how bottom-
up and top-down attentional mechanisms contribute to
successful encoding. In order to do this, we employed
a novel variant of the DM paradigm that was neverthe-
less reminiscent of early manipulations of item presen-
tation rate during learning (Glanzer & Cunitz, 1966).
In this paradigm, we varied the time available prior to
stimulus presentation independently of the time subse-
quent to stimulus presentation. In the encoding phase
of the experiment, subjects viewed a visual event every
2000 mseg, but only one third of these events were
study items (which they were instructed to encode);
the remaining items were fixation crosses. A sequence
of events was constructed such that an equal number
of the possible permutations of zero, uno, or two fixa-
tion cross-stimuli preceding and following the stimulus
was presented in each block. De término medio, de este modo, cada
second visual event was a study item, but crucially,
subjects’ expectations varied in an ‘‘aging interval’’
fashion following the preceding study item. Porque
there were never more than two crosshairs between
elementos, study items that were preceded by two cross-
hairs were predictable because subjects could be 100%
certain that a to-be-encoded stimulus would be pre-
enviado. Por el contrario, immediately following each study
item, there was only a 33% chance that another study
item would be presented, so these items were un-
predictable. We reasoned on the predictable trials,
subjects would be able to orient their attention (en un
top-down fashion) towards the current stimulus, leading
to improvement in memory performance. Por el contrario,
on those trials that were unpredictable, we hypothe-
sized that neural correlates of bottom-up attentional
Summerfield and Mangels
1121
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
orienting would turn out to be robust predictors of
subsequent memory.
In order to explore neural correlates of top-down and
bottom-up attention that predicted subsequent memory
during encoding, scalp EEG activity was recorded during
the study phase of the experiment. We used signal
processing techniques that involve decomposing the
EEG signal
into its spectral components in order to
explore how synchronous neural activity (4–55 Hz)
varied with expectation and learning. En particular, nosotros
were interested in how rhythmic EEG activity at 4–8 Hz
(theta-band activity [TBA]) and at 25–55 Hz (gamma-
band activity [GBA]) varied with manipulations of atten-
tion and encoding. TBA over the dorsomedial frontal
corteza (‘‘frontal midline theta’’ or ‘‘FM theta’’) is a cor-
relate of successful encoding (Summerfield & Mangels,
2005a; Weiss & Rappelsberger, 2000; Klimesch et al.,
1996) and working memory processes (Rizzuto et al.,
2003; Raghavachari et al., 2001; Gevins & Herrero, 2000;
Sarnthein, Petsche, Rappelsberger, Shaw, & von Stein,
1998). Recientemente, reset of the phase of TBA by bottom-up
stimulation has been linked to the activation of atten-
tional control processes (Makeig, Delorme, et al., 2004)
giving rise to a family of ERP components known to
underlie attentional orienting and encoding (como el
P2 and P300). We thus conjectured that TBA might
contribute to encoding as it reflects the bottom-up
activation of attentional control mechanisms triggered
by novel or unpredictable stimulation (such as where
one study item followed unexpectedly on from another
study item).
Recent reports have indicated that >25 Hz activity
(GBA) may also predict later memory (Gruber, Tsivilis,
et al., 2004; Sederberg et al., 2003). This finding coin-
cided with the emergence of a prominent theory link-
ing high-frequency oscillatory activity in the early stages
of the poststimulus period ((cid:1)150 mseg) to top-down
attentional processes (Herrmann, Munk, & ángel, 2004;
Debener, Herrmann, Kranczioch, Gembris, & ángel, 2003;
ángel, Fries, & Cantante, 2001) or expectation (Miltner,
Braun, arnold, Witte, & Taub, 1999). De este modo, one possibil-
ity is that the involvement of neocortical GBA in suc-
cessful encoding reflects top-down attentional processes
that facilitate orienting of attention to an expected or
predictable study item rather than declarative memory
trace formation per se. We thus hypothesized that GBA
would predict later memory for predictable trials, cual
were preceded by a sufficiently long prestimulus pe-
riod for attention to be oriented to them in a predictive
moda.
In a previous study from our laboratory with a similar
diseño, subjects learned word–color associations with a
fixed interval between study items (Summerfield &
Mangels, 2005a). en este estudio, theta-band synchrony
proved to be a particularly reliable predictor of the en-
coding of the association between the word and the
color rather than of later recognition memory for the
word alone. Drawing upon this finding, in the present
study we chose to define later memory for the word–
color association as criterial
for successful episodic
encoding. This decision also reflected the consideration
that contributors to the variance in encoding success
that are tapped by the DM paradigm, such as attention,
do not impact encoding monolithically. Por ejemplo,
the impairment to later memory under divided attention
is more manifest when the criterion for later retrieval is
vivid recollective details of the learning episode, com-
pared to where subjects are merely required to recog-
nize an item as old or new on the basis of a sense of
semantic ‘‘gist’’ or familiarity (Gardiner & Parkin, 1990).
Although it is thought that the mechanisms by which
new associations are encoded overlap with those lead-
ing to later conscious recollection (Hockley & Consoli,
1999), it remains controversial whether divided atten-
tion has greater effect on associative encoding than
on item-only encoding (Castel & Craik, 2003; Naveh-
Benjamín, Guez, et al., 2003). Our study was not in-
tended to speak specifically to this debate, but rather to
use word–color encoding as a model for how a new
episodic memory trace, consisting of composite sensory
and semantic information, is formed under varying con-
ditions of bottom-up and top-down attention.
MÉTODOS
Subjects
Nineteen paid volunteers (12 women) were recruited via
posters placed in Columbia University’s psychology de-
partment. Subjects were neurologically normal right-
handers aged between 18 y 35 años. All subjects gave
informed consent to participate in the study, which was
approved by Columbia University Institutional Review
Board.
Procedimiento
Four study-test blocks were presented. In each study
phase, subjects viewed lists of 45 medium-frequency
(length 3–11 letters) nouns in one of four font colors:
rojo, yellow, green or blue. Words remained on the
screen for 1000 msec and were followed by a blank
screen for 1000 mseg. Interposed between each word
were zero, uno, or two successively and centrally pre-
sented crosshair stimuli. Crosshairs also remained on
the screen for 1000 msec and were followed by a blank
screen for 1000 mseg. Stimulus sequences were carefully
generated such that in each block (de 45 elementos) each of
the nine possible permutations of crosshairs before and
after the study item was presented an equal number of
veces (permitting, across the four blocks, 20 trials per
level of pre and post; for EEG analyses, 60 trials per level
of pre collapsed across post). Filler words were pre-
sented at the beginning and end of each block. Memoria
1122
Revista de neurociencia cognitiva
Volumen 18, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
performance for these filler words was not examined.
Subjects were instructed to ignore the crosshairs and to
remember as many of the 45 word–color associations as
posible. Although no training was given, subjects were
verbally instructed that no more than two crosshairs
would be presented between words.
Following a brief distracter task (counting backwards
by threes from a random three-digit number for 20 segundo),
subjects undertook the test/retrieval phase,
en el cual
they were presented with 90 probe words in white font,
half of which were old (es decir., had been shown in the
immediately prior encoding phase) and half of which
were new (distracters). Two thousand milliseconds after
presentation of the word, subjects were prompted to
make a keypress to indicate whether the probe was old
or new, and to assess their confidence in this judgment
by responding in one of four categories: ‘‘sure old,''
‘‘think old,’’ ‘‘think new,’’ or ‘‘sure new.’’ Immediately
following this response, a second prompt appeared,
asking subjects to indicate in which of the four font
colors the word had been presented. De nuevo, for each
option, subjects could indicate that they were ‘‘sure’’ or
that they ‘‘guessed’’ that this was correct, giving a total
of eight response options (‘‘sure red,''
‘‘guess red,''
‘‘sure blue,’’ etc.). Following this response, a blank
screen was presented for 1000 mseg, and then the next
memory probe.
EEG Recording and Artifact Rejection
EEG activity was recorded during the encoding and
retrieval phases of the experiment, although in this
report only data from the encoding phase are discussed.
EEG was acquired from 64 canales (sampling rate =
500 Hz; high-pass filter = 0.1 Hz,
low-pass filter =
100 Hz; impedances kept below 11 k(cid:1)) using Neuro-
scan SYNAMPS (Compumedics Inc., El Paso, Texas). Re-
cordings were initially referenced to Cz, then converted
to an average reference off-line. BESA 5.06 (Electrical
Geodesics Inc., Eugene, O) was used to remove eye
movimientos, parpadea, and other artifacts from the con-
tinuous EEG data (Iceberg & Scherg, 1994). Trials in which
muscle, movimiento, or other artifacts drove amplitude
values above 100 AV were manually rejected. Artifact-free
data were high-pass filtered with a 0.5-Hz zero-phase
filter, spline-Laplacian transformed (estimated dura po-
tential) (Nunez et al., 1997), and interpolated into an
81-electrode montage. Epochs from 1000 msec prestim-
ulus to 2000 msec poststimulus then were exported into
ASCII format.
Data Analysis with Brain Electrophysiology
Analysis and Statistical Testing
All further data processing was performed with in-house
MATLAB software written by CS. The code used for
signal processing and analysis (brain electrophysiology
analysis and statistical testing [BEAST]) may be freely
downloaded at www.columbia.edu/~cs2028/ beast/
beast.htm. A detailed explanation of the analysis tech-
nique has been described elsewhere (Summerfield &
Mangels, 2005a). Under the framework of this analysis
package, wavelet-transformed data are subjected to in-
dependent components analysis (ICA) to separate the
data into spatially correlated sources of variance. Signif-
icance testing is carried out using random permutation
testing on component activation scores, and compo-
nent projections are visualized on a 2-D scalp map to
show brain regions contributing maximally to the com-
ponent activations. BEAST uses ICA code from EEGLAB
(Delorme & Makeig, 2004).
Wavelet Power
All epochs were Morlet wavelet-transformed (Torrence,
& Campo, 1998) entre 4 y 13 Hz (theta/alpha
analiza) y 14 y 55 Hz (beta/gamma analyses) en
nine logarithmically spaced frequency bands (vo = 6).
To reduce artifacts, wavelet values were windsorized at 5
standard deviations above the mean. Data were tempo-
rally smoothed with a Gaussian filter of width 50 mseg,
and averaged across adjacent frequency bands, to in-
crease signal/noise ratio. Trials in each condition were
subsequently averaged, yielding mean time–frequency
(TF) information for each condition, at each electrode,
for each subject. Averaged wavelet values were con-
verted to z scores at each subject and electrode prior
to ICA analyses and significance testing.
ICA and Significance Testing
Data were reduced to correlated spatial components
with ICA, and significance testing was carried out at each
component with nonparametric random permutation
pruebas. Analyses of variance (ANOVAs) with planned
comparisons were carried out in parallel at every TF
pixel to derive point-estimate statistics, and then subject-
condition values were shuffled 1000 times and this
ANOVA repeated for each permutation. To provide a
correction for ‘‘familywise error’’ multiple comparisons,
the maximum value across the entire TF plot was then
entered into a distribution that forms the null against
which point estimate values are compared. Only those
point estimates that fall within the top 5% of this
distribution were considered significant. This technique
has been described in more detail elsewhere (Burgess &
Gruzelier, 1999).
The present factorial experimental design presented a
unique challenge for this method, as the interaction
term statistics cannot be reliably calculated using the
method described above (Suckling & bullmore, 2004).
We thus used planned comparisons to derive regions of
interés (ROI) within TF space, and then tested mean
wavelet power/coherence values falling within this ROI
Summerfield and Mangels
1123
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
using conventional parametric statistics. The planned
comparisons used for mem and pre searched for voxels
that varied for WC > miss or miss > WC, collapsed
across pre; for pre, the search was for voxels that varied
in a pre2 > pre1 > pre0 or pre0 > pre1 > pre2 fashion,
collapsed across mem. For the Pre (cid:2) Mem interaction,
we searched for voxels that predicted later memory only
at pre0, pre1, or pre2 separately, or that varied across
pre for WC or miss trials separately. Because planned
comparisons are mean driven,
en algunos casos, vóxeles
satisfied these conditions but did not achieve signifi-
cance when the interaction term was calculated with
conventional parametric statistics. These effects are not
reported here.
RESULTADOS
Behavioral Results
Study-phase trials were classified on the basis of whether
subjects, on a later retrieval test, (a) correctly judged the
item to be old and correctly identified in which font
color it had been presented (‘‘word + color’’ or WC
ensayos) o (b) either failed to correctly recognize the item,
or recognized the item but failed to correctly recollect
the font color in which it was presented (‘‘miss’’ trials).
In order to increase statistical power, we collapsed
across high- and low-confidence responses for all analy-
ses. Three subjects were excluded, as they failed to
identify the font color with above-chance accuracy (leav-
ing n = 16). En general, font color was correctly recollected
on 42–86% of trials (mean = 64 ± 16%; chance = 25%)
in the remaining subjects.
In order to investigate how memory performance var-
ied with prestimulus time (pre) and poststimulus time
(post) tiempo, trials were labeled according to the number
of crosshairs preceding and following the stimulus; para
ejemplo, in the stimulus sequence A B + + C, dónde
stimulus B followed directly on from stimulus A, pero
two crosshairs intervened between stimuli B and C,
stimulus B would be classified as ‘‘pre0’’ and ‘‘post2.’’
A schematic diagram of the trial sequence can be seen
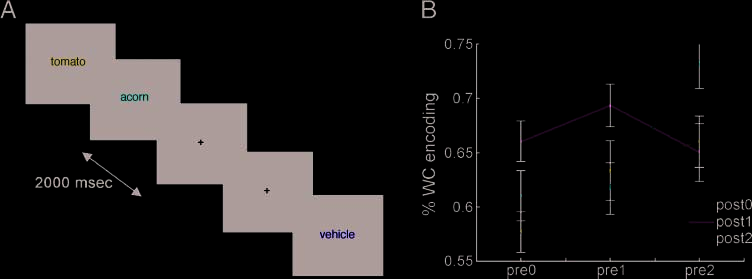
in Figure 1A. Figure 1B shows memory performance
(percent word–color associations learned) as a function
of pre- and poststimulus time. Visual inspection shows
an overall pattern of increasing memory performance
with increasing time preceding the stimulus. En efecto,
when memory performance was compared using facto-
rial analysis of variance (pre [3] (cid:2) post [3]), later mem-
ory for the word–color association varied reliably with
pre (F = 5.59, pag < .01) in a pre2 > pre1 > pre0 fashion
(linear trend: F = 9.0, pag < .01). A significant main effect
of post (F = 8.65, p < .01) was also observed, again with
later memory performance increasingly linearly across
poststimulus time (F = 13.8, p < .01). No interaction
between pre and post was found.
EEG Results
In order to explore how the memory advantage for
predictable trials effect might be reflected in brain
activity accompanying the study phase, we examined
how neural synchrony associated with encoding trials
varied as a function of both subsequent memory (mem)
and expectation (pre). In order to provide full coverage
of the EEG frequency spectrum, power information was
measured in the theta/alpha (4–13 Hz) and beta/gamma
(14–55 Hz) bands for an epoch lasting for 800 msec
following stimulus presentation. ICA of gamma- and
theta-band data were performed separately to facilitate
the isolation of unrelated sources of variance in each
frequency band. All EEG data presented here are from
the encoding phase of the experiment, and only re-
sponses to study items (not to interspersed crosshairs)
are shown.
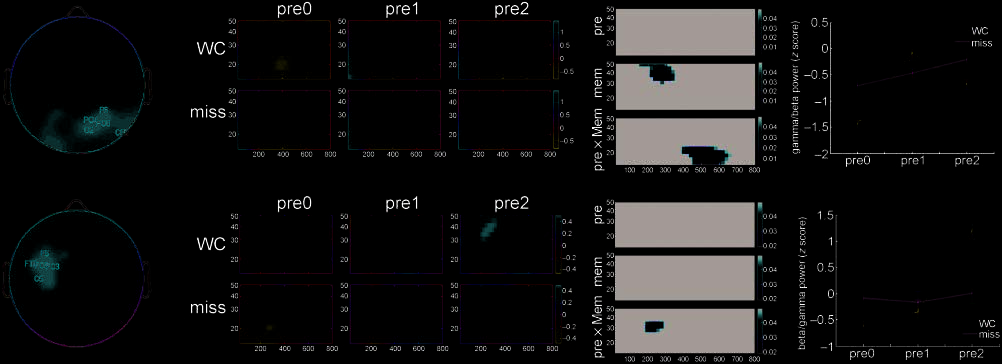
Graphical displays of spectral power results (Figures 2
and 3) each consist of four panels: (i) Component
weights are plotted on the scalp to show the spatial
Figure 1. Encoding phase
trial sequence and behavioral
data. (A) Subjects were
presented with a visual event
every 2 sec, one third of which
was a study item (word in
one of four font colors). Prior
to and following each study
item, zero, one, or two fixation
crosshairs were presented.
Study items were classified
according to the number
of crosses preceding and
following the stimulus: For
example, the word ‘‘acorn’’
here is preceded by 0 and
followed by two crosshairs (pre0/post2). (B) Behavioral performance. On the y-axis, the percentage of trials on which the word–color
association was subsequently remembered for each level of pre and post. On the x-axis, pre0 (left), pre1 (middle) and pre2 (right) represent
trials preceded by zero, one, and two crosshairs, respectively. Post0, post1, and post2 are plotted with blue, green, and red lines, respectively.
1124
Journal of Cognitive Neuroscience
Volume 18, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
7
-
1
p
1
d
2
f
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
o
0
c
/
n
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
o
1
c
2
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
7
t
.
o
1
n
1
0
2
7
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. EEG results: 4–13 Hz. Left: component weights for each electrode. Middle left: TF plots (x-axis: time, 0–800 msec; y-axis:
frequency, 4–13 Hz) for trials on which the word–color association was successfully learned ( WC) and for which it was not (miss) for study
trials preceded by zero (pre0), one (pre1), or two (pre2) crosshairs. Values are mean z scores across subjects. Middle right: significance
plots for the main effect of pre (top), the main effect of mem (middle) and the Pre (cid:2) Mem interaction (bottom). Only pixels significant at
p < .05 (corrected for multiple comparisons) are shown; all other pixels are black. Right: mean spectral power values extracted from a mask
in TF space corresponding to pixels exceeding p < .05 for the main effect of mem at Component 1 (top) and Mem (cid:2) Pre at Component 2
(bottom). Mean spectral power across subjects is plotted on the y-axis; pre0, pre1, and pre2 are on the x-axis. The blue line represents
WC trials, and the green line represents miss trials.
distribution of each component (leftmost); (ii) mean
component activations (TF plots, averaged across sub-
jects) are shown for each level of pre and mem (middle
left); (iii) planned comparisons (with appropriate cor-
rection for multiple comparisons) were used to isolate
regions of TF space where activation scores for each
component differed reliably as a function of pre, mem
and Pre (cid:2) Mem (middle right); and (iv) these regions
were then used as masks to extract mean spectral power
for each level of pre and mem from the component
activation plots, and these effects are then displayed on
separate axes (rightmost).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
8
e
7
-
1
p
1
d
2
f
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
o
0
c
/
n
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
o
1
c
2
n
0
.
p
2
d
0
0
b
6
y
.
g
1
u
8
e
.
s
7
t
.
o
1
n
1
0
2
7
0
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
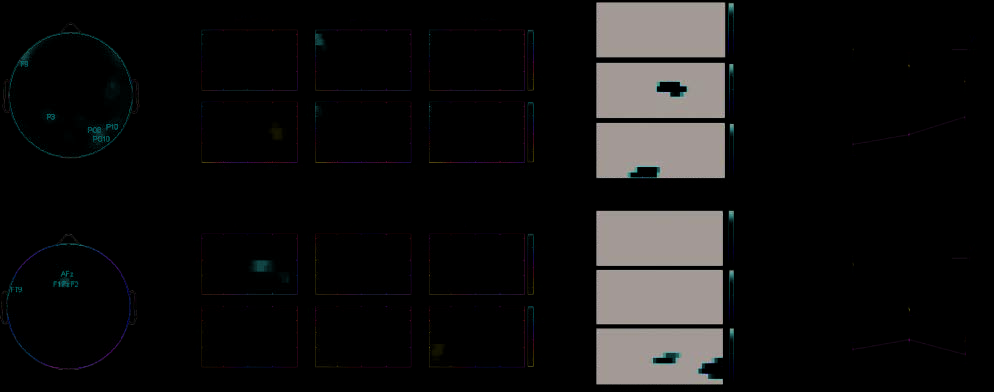
Figure 3. EEG results: 14–55 Hz. Left: component weights for each electrode. Middle left: TF plots (x-axis: time, 0–800 msec; y-axis:
frequency, 14–55 Hz) for trials on which the word–color association was successfully learned (WC) and for which it was not (miss) for study
trials preceded by zero (pre0), one (pre1), or two (pre2) crosshairs. Values are mean z scores across subjects. Middle right: significance plots
for the main effect of pre (top), the main effect of mem (middle), and the Pre (cid:2) Mem interaction (bottom). Only pixels significant at p < .05
(corrected for multiple comparisons) are shown; all other pixels are black. Right: mean spectral power values extracted from a mask in TF
space corresponding to pixels exceeding p < .05 for the main effect of Pre (cid:2) Mem at Component 1 (top) and Mem (cid:2) Pre at Component 2
(bottom). Mean spectral power across subjects is plotted on the y-axis; pre0, pre1, and pre2 are on the x-axis. The blue line represents WC
trials, and the green line represents miss trials.
Summerfield and Mangels
1125
EEG Results: 4–13 Hz Data
Characteristic EEG responses followed each visual event
in the encoding phase of the experiment. Across the
entire scalp, stimulus presentation triggered a steady
slowing of the EEG response, leading to a characteristic
diagonal ‘‘alpha–theta shift’’ in the TF plots throughout
the first 400 msec of the epoch (Figure 2). Although this
pattern of neural activity was widespread across the
scalp, highest ICA weights were observed over the left
(P3) and right (PO8, PO10, P10) parietal cortex, as well
as over left frontal sites (F9). From (cid:1)400 msec onward,
4- to 13-Hz EEG responses at this component de-
synchronized, but deeper theta/alpha desynchronization
was observed for miss trials, leading to a subsequent
memory effect in the theta-band at 400–600 msec post-
stimulus (F = 12.7, p < .003). These effects were
described by an ICA component that explained 28.6%
of the variance in poststimulus theta/alpha-band activity
(Figure 2, top).
A second component (variance explained = 4.7%)
loaded on anterior midline electrodes (AFz, Fz, F1, F2)
extending more widely over left frontal regions ventro-
laterally as far as anterior temporal electrode FT9. At
these electrodes, TBA (<8 Hz) onset was at (cid:1)300 msec
and continued until the end of epoch, and neural
synchrony>400 msec predicted later memory only
for those study items that were immediately preceded
by another study item (pre0 trials) (Pre (cid:2) Mem interac-
ción, F = 3.4, pag < .05). A main effect of mem (F = 9.7,
p < .008) was also observed for this late TBA, and an
effect of pre approached significance (F = 2.6, p < .09).
Results for this component can be seen in Figure 2
(bottom).
EEG Results: 14–55 Hz Data
Results for the gamma/beta frequency ranges are shown
in Figure 3. The most characteristic feature of rhythmic
EEG activity in the beta/gamma ranges was a powerful
desychronization following stimulus presentation, which
was maximal over right parieto-occipital electrodes
(O2, CB2, PO4, PO8, and P6). This desynchronization
succeeded a burst of beta/alpha power concurrent with
stimulus presentation, which persisted in the lower beta
range until (cid:1)100 msec poststimulus. Encoding success
modulated this response in the higher frequency range,
as for WC trials, the 40–50 Hz response tapered off
very early in the epoch, but for miss trials, it continued
until >300 msec poststimulus. This led to reliable dif-
ferences in 40–50 Hz activity as a function of later mem-
ory, with reduced neural synchrony over posterior scalp
sites at >150 msec observed for later remembered trials
(F = 8.0, pag < .02). This effect can be seen in the second
from the top significance plot (‘mem’) in Figure 3 (top,
middle right panel). No main effect of pre ( p = .22) or
interaction ( p = .54) were observed within this mask.
However, later in the epoch, beta-band activity varied
in an interacting fashion with subsequent memory and
anticipation time, with a deeper desynchronization pre-
dicting later memory only for pre0 trials (F = 6.3, p <
.01) (Figure 3, bottom right). This effect was reliable
from 400 to 600 msec, at 14–25 Hz (Figure 3, bottom,
middle right). This ICA component explained 13.1% of
the variance in poststimulus beta/gamma activity.
A second component (variance explained = 12.2%)
exhibited a broad frontotemporal topography, with
maximal weights observed at electrodes F5, FT7, FC5,
C3, and C5. Neural activity onset over these regions
in the beta/gamma range with or shortly after stimulus
presentation, and frontal (cid:1)40 Hz synchrony from (cid:1)200–
300 msec predicted subsequent memory, but only in
the pre2 condition, where the study item was preceded
by two crosshair stimuli. This effect was statistically
reliable (Pre (cid:2) Mem interaction, F = 4.43, p < .03)
although the focus of the statistical differences was
slightly lower, at 30–35 Hz. No main effect of mem
( p = .68) was observed within this mask over these
frontal sites, although there was a trend for GBA to
increase in concert with anticipation time ( p = .06).
DISCUSSION
Subjects performed a version of the DM paradigm in
which the times that preceded and followed each study
item were independently manipulated. As predicted,
a longer poststimulus period facilitated later memory,
presumably because it permitted more time for strate-
gic conceptual processing (such as phonological rehears-
al or semantic elaboration) that is well known to lead
to ‘‘deep’’ encoding (Craik & Lockhart, 1972). How-
ever, the novel behavioral result presented here is that
time prior to the stimulus is a robust and independent
predictor of encoding success, with longer intervals
prior to presentation of a study item leading to im-
proved later memory for that item. Moreover, we iden-
tified dissociable subsequent memory effects over the
left prefrontal cortex for predictable study items (which
subjects were 100% sure were about to occur) and
unpredictable study items (which there was only a
33% chance would occur) in the gamma (>25 Hz) y
theta (4–8 Hz) bands, respectivamente. These findings com-
plement previous research into the functional role
of gamma- and theta-band neural synchrony in mem-
ory and attention, and describe,
por primera vez,
functionally dissociable correlates of top-down and
bottom-up attention orienting during successful epi-
sodic encoding.
Neural Synchrony: Theta Band (4–8 Hz)
Compelling evidence indicates that TBA plays a role in
short-term memory maintenance (Sarnthein et al., 1998)
1126
Revista de neurociencia cognitiva
Volumen 18, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
F
.
.
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
but the precise nature of that role remains poorly
comprendido (Bastiaansen & Hagoort, 2003). Otro
role in intermediate- and long-term memory formation is
suggested by the finding that long-term potentiation,
the candidate cellular–molecular basis for the formation
of new memories, seems to depend on the phase of
hippocampal theta activity (Huerta & Lisman, 1993), y
blocking theta activity by lesioning the medial septum
causes spatial memory impairments (Givens & Olton,
1990). Además, during serial
en-
creases in neocortical theta-band synchrony, particularly
over the frontal cortex, are a robust predictor of later
memory (Summerfield & Mangels, 2005a; Montaña de cedro
et al., 2003; Klimesch et al., 1996). Neocortical TBA
may thus play an important role in memory formation,
perhaps by ‘‘informing’’ the hippocampus what should
be bound with what during declarative memory forma-
ción (Buzsaki, 1996).
learning of words,
Sin embargo, a broader role for TBA in cognition has
been suggested by the finding that sensorimotor and
attentional mechanisms are associated with increases in
theta-band synchrony. En particular, frontal midline TBA
seems to reflect control processes that are activated
when a subject is required to orient attention or prepare
to respond (Makeig, Delorme, et al., 2004), and ERP
components implicated in detection of novel or salient
events, such as the positive-going P2 and P300 waves ob-
served over midline frontal and parietal electrodes, may
reflect event-related spectral perturbation (ERSP) of on-
going theta rhythms at the cortex (Makeig, Westerfield,
et al., 2002). According to this theory, bottom-up stim-
ulation may lead to a ‘‘phase reset’’ of the theta rhythm,
such that during event-related averaging, amplitude
maxima and minima of ongoing theta activity are aligned
to form the characteristic positive- and negative-going
deflections of the ERP (although this view of the rela-
tionship between neural synchrony and the ERP remains
controversial) ( Yeung, Hombre rico, Holroyd, & cohen, 2004).
Aquí, we argue that a theta-band phase-reset may form
part of a mechanism by which attention is oriented to
an unpredicted stimulus,
invoking executive control
processes required to flexibly accommodate novel or
unexpected information within the current set of goals
and plans. Respectivamente, decades of ERP research have
shown that positive-going deflections at 200–400 msec
over the frontal and parietal cortices are enhanced
during the processing of oddball stimuli (McCarthy &
Donchin, 1981), are suppressed under divided atten-
ción (Mangels et al., 2001; Wickens, Kramer, Vanasse, &
Donchin, 1983), and predict subsequent memory (Otten
& Donchin, 2000), precisely as would be predicted if
these early ERP/ERSP effects reflect attentional effects
that gate successful encoding. The idea that frontal
midline TBA reflects a general attentional orienting
mechanism can explain why it has been found to vary
over a wide range of attentional, memory, and motoric
tareas, and why abnormalities in TBA are a hallmark of
disorders of attention such as ADHD (Barry, Clarke, &
Johnstone, 2003).
Regardless of whether the stimulus was predictable
(pre2) or not (pre0), we observed increases in TBA all
over the scalp, and notably over parietal electrodes, en
trials on which the word–color association was success-
fully encoded (WC trials) compared to those on which it
was not (misses). We thus replicated previous reports
implicating TBA in successful encoding (Summerfield &
Mangels, 2005a; Sederberg et al., 2003; Klimesch et al.,
1996). More importantly, sin embargo, both at this broad
scalp component and at another component that loaded
heavily on midline frontal scalp sites and extended over
the left frontal cortex, a significant interaction was ob-
served whereby theta synchrony predicted later mem-
ory to a greater extent on pre0 trials, eso es, dónde
one study item unexpectedly succeeded another. Este
activity was initially observed as early as (cid:1)200 mseg
poststimulus at posterior electrodes, and slightly later
(>400 msec) over the frontal cortex, falling well within
the time frame of observed theta-band ERSP effects. Él
seems likely that where the appearance of a forthcom-
ing stimulus cannot be predicted, the effectiveness with
which attention is disengaged from the prior study item
and oriented toward the current item will be a robust
predictor of subsequent memory. Por el contrario, on well-
spaced, predictable trials, TBA is a poorer predictor of
later memory, as subjects are able to orient their at-
tention to the forthcoming study item in a top-down
fashion even before the stimulus arrives. Además, el
topography of the ICA component at which this activity
loading maximally on frontal midline
was observed,
electrodes,
is highly consistent with previous studies
showing that frontal dorsomedial areas (and the under-
lying anterior cingulate cortex) are an important part
of a network underlying the orienting of attention to
spatial locations, características, or representations (Corbetta
& Shulman, 2002). These data thus provide support for
the idea that TBA predicts later memory because it sub-
serves the disengagement and reengagement of atten-
tion during the unfolding of an unpredictable sequence
of stimuli or events.
Neural Synchrony: Gamma Band (>25 Hz)
We also found increases in GBA over the left prefrontal
cortex during successful encoding. This is consistent
with recent reports that increased neocortical gamma-
band synchrony during encoding is associated with later
recordar (Sederberg et al., 2003) and recognition memory
(Gruber, Tsivilis, et al., 2004). Does neocortical GBA
directly contribute to the formation of a new declarative
memory trace, or does it covary with attentional factors
that facilitate encoding? The former is suggested by data
from intracranial recordings in humans, which noted
that phase-locking of gamma-band responses between
the hippocampus and nearby rhinal cortex (Fell, Klaver,
Summerfield and Mangels
1127
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
F
.
.
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Lehnertz, et al., 2001), through which major afferent
pathways to the hippocampus pass (Amaral & Witter,
1989). Sin embargo, accumulating evidence suggests a role
for neocortical GBA in the top-down orienting of atten-
tion to an expected or target stimulus (Herrmann et al.,
2004; Debener et al., 2003; Engel et al., 2001). Consist-
ent with this view, we observed that on trials where the
presentation of a study item was predictable (es decir., cuando
subjects had time to orient attention in a top-down
fashion to a forthcoming event) el (cid:1)40 Hz response
was earlier and stronger as a function of later memory.
The functional significance of high-frequency brain
actividad (>25 Hz) in human cognition remains contro-
versial (Kaiser & Lutzenberger, 2005). Early reports ar-
guing for a role in bottom-up perceptual grouping
according to gestalt principles (Cantante & Gray, 1995)
were quickly complemented with findings that GBA is
observed to vary with experimental manipulations of
atención ( Muller, Gruber, & Keil, 2000; Gruber, Muller,
Keil, & Elbert, 1999; Tallon-Baudry, Bertrand, Delpuech,
& Permier, 1997) and working memory (Tallon-Baudry,
Bertrand, & pescador, 2001). Respectivamente, a role in top-
down attentional control has been proposed (ángel
et al., 2001). According to one model, GBA may be in-
volved in ‘‘matching’’ a top-down prediction to bottom-
up sensory information in order to facilitate recognition
of an expected (Miltner et al., 1999) or relevant (Debener
et al., 2003; Fries, Reynolds, Rorie, & Desimone, 2001)
stimulus once it arrives. This model has been called
memory match and utilization (MUM) theory (Herrmann
et al., 2004) and argues that ‘‘early’’ gamma activity
(occurring at (cid:1)150 msec poststimulus) reflects the con-
fluence of incoming information with a top-down repre-
sentation or preestablished schema. Intuitivamente, it makes
sense that such a mechanism should be deployed where
the to-be-learned stimuli are expected and a reliable
top-down prediction can be formed of the forthcoming
stimulus (Friston, 2003), as was the case on pre2 (‘‘pre-
dictable’’) ensayos. Furthermore the time and frequency at
which we observed subsequent memory effects (40 Hz,
(cid:1)200 msec poststimulus) show very close convergence
with those conjectured to underlie the confluence of
top-down and bottom-up activity. Our data are thus
highly consistent with the MUM model, and extend it to
suggest that in addition to facilitating recognition of the
stimulus, these top-down attentional mechanisms may
increase the likelihood that it is encoded into long-
term memory. Curiosamente, a recent study of sentence
comprehension has shown that GBA predominates when
sentences contain semantically correct or expected
elementos, whereas theta-band responses are evoked by se-
mantic violations, data that clearly mimic the theta/
gamma dissociation reported here (Hald, Bastiaansen,
& Hagoort, 2005).
Over posterior electrodes, sin embargo, a different pat-
tern was observed whereby gamma-band synchroniza-
tion at 40–50 Hz was reliably greater during unsuccessful
encoding, an effect that occurred at (cid:1)300 msec post-
stimulus. Although it may at first appear contradictory,
‘‘negative’’ subsequent memory effects (es decir., desynchro-
nization associated with successful encoding) ha sido
reported previously in the gamma band (Montaña de cedro
et al., 2003; Fell, Klaver, Lehnertz, et al., 2001). En efecto,
recordings from within the hippocampus have shown
that enhanced gamma-band spectral power at 100–
400 msec is a correlate of encoding trials that are later
‘‘missed,’’ a time course that closely matches that ob-
served here.
Left Anterior Cortex and Episodic Encoding
Neural activity that varied in an interacting fashion be-
tween both top-down and bottom-up attentional orient-
ing and episodic encoding extended over scalp electrodes
that were positioned to capture neural activity from the
left prefrontal cortex. Left dorsolateral prefrontal regions
have been previously shown to be a robust predictor
of the learning of word–color associations (Ranganath
et al., 2004), and more ventral regions are among the
most frequently implicated in subsequent memory stud-
ies where item-context binding is criterial for successful
encoding (Davachi, mitchell, & Wagner, 2003; Cansino,
Maquet, Dolan, & Rugg, 2002). Además, activity in
the left prefrontal cortex fails to predict later memory
when attention is divided with a demanding secondary
tarea, indicating that the contribution of this region to
successful encoding is attention dependent. This result,
coupled with the established theory that left frontal
regions support semantic retrieval (Cabeza & Nyberg,
2000), and the finding that it is recall and conscious
recollection (retrieval criteria that benefit most from
‘‘deep’’ encoding) that suffer most under divided atten-
ción (Craik et al., 1996), forms the basis for the view that
reducing attentional resources impairs later memory be-
cause it leaves subjects less opportunity to engage in
deep semantic elaboration of the study item.
Sin embargo, it has also been suggested that suppression
of semantic elaboration may not be sufficient to account
for the impairment to later memory observed following
divided attention (Naveh-Benjamin, Guez, et al., 2003).
Por ejemplo, DA-related impairments in later recall
persist even when items were matched with respect to
subjective ratings of how well they were elaborated at
estudiar (Craik & kester, 2000). One explanation for this
discrepancy is that over and above any role in semantic
retrieval, left anterior regions may form part of a wider
network that contributes to successful encoding by me-
diating the control of attention toward the study item.
A more direct role for the left anterior cortex in atten-
tion processing has been suggested by a previous DM
study that found ‘‘state-related’’ effects predicting later
memory in the left inferior frontal lobe (Otten, Henson,
& Rugg, 2002). State-related effects index neural activ-
ity that does not vary with the temporal onset of the
1128
Revista de neurociencia cognitiva
Volumen 18, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
F
.
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
stimulus, such as slow fluctuations in vigilance, or atten-
tional control processes that begin prior to stimulus
presentación. This study, taken together with others
positing a direct role for the left anterior cortex in the
controlled biasing of attention (Brass & por cramon,
2004; macdonald, cohen, Stenger, & Carretero, 2000),
offers an alternative explanation for our data: that the
contribution of left frontal regions to episodic encod-
ing relates to controlled orienting of attention to pre-
dictable and unpredictable verbal events. This view is
consistent with recent approaches to understanding
the function of the left prefrontal cortex that eschew
domain specificity in favor of a wider role in the reg-
ulation of mental activity (Thompson-Schill, Bedny, &
Goldberg, 2005).
Neural Synchrony: Other Results
Although we observed increases in gamma-band syn-
chronization over left frontal regions in association with
later memory, the general trend for high-frequency
(>14 Hz) neural activity across more posterior cortical
regions was to desynchronize upon stimulus presenta-
ción. Neural activity in the beta and gamma bands
exhibited a characteristic prestimulus synchronization
that tapered off earlier at higher frequencies, continuing
in the lower beta band until about (cid:1)100 msec poststim-
ulus, but then gave way to a powerful desynchronization
that was prominent across the scalp. ICA analyses in our
laboratory (Summerfield & Mangels, 2005b) y estafa-
ventional analyses from other laboratories (Bastiaansen,
van der Linden, Ter Keurs, Dijkstra, & Hagoort, 2005)
have previously identified this trend in the beta fre-
quency range, although it is most often observed fol-
lowing a voluntary movement (Pfurtscheller, 1992). El
extent of beta synchrony varied with expectation and
memory, with a deeper desynchronization on pre0/ WC
ensayos, where subjects successfully learned the second
of two sequentially presented word–color associations.
Beta desynchronization at >500 msec has been previ-
ously observed in association with subsequent memory
(Sederberg et al., 2003), a pesar de, en general, little is
known about its role in attention and memory. Cómo-
alguna vez, a recent report employing rapid serial visual pre-
sentation has argued that beta-band synchronization
may reflect a state of increased vigilance for behaviorally
relevant stimuli (Gross et al., 2004). It could be, de este modo,
that in conditions where bottom-up attention is not
captured by the stimulus (es decir., where the study item is
predictable, or where subjects fail to orient and thus
encode), processing resources are freed up for vigilant
anticipation of the forthcoming stimulus. One explana-
tion for our observation of a beta-band ‘‘rebound’’ in a
nonmotoric task is thus that it reflects generalized antic-
ipation processes relating to the forthcoming stimulus,
rather than the neural sequelae of voluntary movement
execution.
Summary
Our data thus argue for a functional dissociation in the
role of theta (4–8 Hz) and gamma (25–55 Hz) activity in
successful episodic encoding. One way of interpreting
the data presented here is that multiple attentional
procesos, dependent on the activation of the frontal
corteza (and in particular left frontal cortex) sites, facilidades-
tate the formation of new memories. Where the to-be-
encoded information is unexpected, the extent to which
attention is disengaged from the current focus and
oriented to the relevant information is likely to be highly
predictive of later memory. This mechanism is mediated
by transient neural synchrony in the theta (4–8 Hz)
banda, particularly over dorsomedial frontal regions pre-
viously implicated in endogenous attentional orienting.
Predictable information, sin embargo, is encoded in a dif-
ferent way. Top-down attentional mechanisms mediated
by neural synchrony in the gamma (>25 Hz) banda
increase early in the poststimulus epoch as a function
of expectancy and memory, in concert with increasing
behavioral performance on trials where study items
were well spaced and expected. Left frontal GBA thus
constitutes an excellent candidate for the neural sub-
strate of a top-down attentional orienting mechanism
that facilitates the encoding of new episodic memories.
Expresiones de gratitud
This research was supported by NIH grant R21 066129 a
j. A. METRO. and a grant from the W. METRO. Keck Foundation to
Columbia University.
Reprint requests should be sent to Christopher Summerfield,
Psychology Department, Schermerhorn Hall, Room 406, Co-
lumbia University, 1190 Amsterdam Ave., Nueva York, Nueva York 10027,
o por correo electrónico: summerfd@psych.columbia.edu.
REFERENCIAS
Amaral, D. GRAMO., & Witter, METRO. PAG. (1989). The three-dimensional
organization of the hippocampal formation: Una revisión de
anatomical data. Neurociencia, 31, 571–591.
anderson, norte. D., Iidaka, T., Cabeza, r., Kapur, S., McIntosh,
A. r., & Craik, F. I. (2000). The effects of divided attention
on encoding- and retrieval-related brain activity: A PET
study of younger and older adults. Revista de Cognitivo
Neurociencia, 12, 775–792.
Barry, R. J., Clarke, A. r., & Johnstone, S. j. (2003).
A review of electrophysiology in attention-deficit/
hyperactivity disorder: I. Qualitative and quantitative
electroencephalography. Clinical Neurophysiology,
114, 171–183.
Bastiaansen, METRO., & Hagoort, PAG. (2003). Event-induced theta
responses as a window on the dynamics of memory.
Corteza, 39, 967–992.
Bastiaansen, METRO. C., van der Linden, METRO., Ter Keurs, METRO.,
Dijkstra, T., & Hagoort, PAG. (2005). Theta responses are
involved in lexical–semantic retrieval during language
Procesando. Revista de neurociencia cognitiva, 17,
530–541.
Summerfield and Mangels
1129
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
F
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Iceberg, PAG., & Scherg, METRO. (1994). A multiple source approach
Fabiani, METRO., Karis, D., & Donchin, mi. (1990). Efectos de
to the correction of eye artifacts. Electroencephalography
and Clinical Neurophysiology, 90, 229–241.
Brass, METRO., & por cramon, D. Y. (2004). Selection for cognitive
control: A functional magnetic resonance imaging study
on the selection of task-relevant information. Diario de
Neurociencia, 24, 8847–8852.
Buckner, R. l., logan, J., Donaldson, D. I., & Rodador, METRO. mi.
(2000). Cognitive neuroscience of episodic memory
encoding. Acta Psicológica, 105, 127–139.
Burgess, A. PAG., & Gruzelier, j. (1999). Methodological advances
in the analysis of event-related desynchronisation data:
Reliability and robust analysis. In G. Pfurtscheller & F. h.
Lopez Da Silva (Editores.), Handbook of electroencephalography
& clinical neurophysiology (páginas. 139–160). Ámsterdam:
Elsevier.
mnemonic strategy manipulation in a Von Restorff paradigm.
Electroencephalography and Clinical Neurophysiology,
75, 22–35.
Fell, J., Klaver, PAG., Elfadil, h., Schaller, C., Elger, C. MI.,
& Fernandez, GRAMO. (2003). Rhinal–hippocampal theta
coherence during declarative memory formation:
Interaction with gamma synchronization? European
Revista de neurociencia, 17, 1082–1088.
Fell, J., Klaver, PAG., Lehnertz, K., Grunwald, T., Schaller, C.,
Elger, C. MI., & Fernandez, GRAMO. (2001). Human memory
formation is accompanied by rhinal–hippocampal coupling
and decoupling. Neurociencia de la naturaleza, 4, 1259–1264.
Fletcher, PAG. C., Shallice, T., & Dolan, R. j. (1998). El
functional roles of prefrontal cortex in episodic memory.
I. Encoding. Cerebro, 121, 1239–1248.
Buzsaki, GRAMO. (1996). The hippocampo-neocortical dialogue.
Fletcher, PAG. C., Shallice, T., & Dolan, R. j. (2000). ‘‘Sculpting
Corteza cerebral, 6, 81–92.
Cabeza, r., & Nyberg, l. (2000). Imaging cognition II: Un
empirical review of 275 PET and fMRI studies. Diario
of Cognitive Neuroscience, 12, 1–47.
Cansino, S., Maquet, PAG., Dolan, R. J., & Rugg, METRO. D. (2002).
Brain activity underlying encoding and retrieval of source
memory. Corteza cerebral, 12, 1048–1056.
Castel, A. D., & Craik, F. I. (2003). The effects of aging and
divided attention on memory for item and associative
información. Psicología y envejecimiento, 18, 873–885.
Corbetta, METRO., & Shulman, GRAMO. l. (2002). Control de
goal-directed and stimulus-driven attention in the brain.
Naturaleza Reseñas Neurociencia, 3, 201–215.
Craik, F., & Lockhart, R. (1972). Levels of processing:
A framework for memory research. Journal of Verbal
Aprendiendo & Comportamiento verbal, 11, 671–684.
Craik, F. I., Govoni, r., Naveh-Benjamin, METRO., & anderson,
norte. D. (1996). The effects of divided attention on
encoding and retrieval processes in human memory.
Revista de Psicología Experimental: General, 125,
159–180.
Craik, F. I., & kester, j. D. (2000). Divided attention and
memory: Impairment of processing or consolidation?
In E. Tulving (Ed.), Memoria, consciousness and the
cerebro: The Tallinn Conference (páginas. 38–51). Filadelfia:
Prensa de Psicología.
Davachi, l., Maril, A., & Wagner, A. D. (2001). Cuando
keeping in mind supports later bringing to mind: Neural
markers of phonological rehearsal predict subsequent
remembering. Revista de neurociencia cognitiva, 13,
1059–1070.
Davachi, l., mitchell, j. PAG., & Wagner, A. D. (2003).
Multiple routes to memory: Distinct medial temporal
lobe processes build item and source memories.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
100, 2157–2162.
Debener, S., Herrmann, C. S., Kranczioch, C., Gembris, D.,
& ángel, A. k. (2003). Top-down attentional processing
enhances auditory evoked gamma band activity.
NeuroReport, 14, 683–686.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Revista de neurociencia
Métodos, 134, 9–21.
ángel, A. K., Fries, PAG., & Cantante, W.. (2001). Dynamic
predicciones: Oscillations and synchrony in top-down
Procesando. Naturaleza Reseñas Neurociencia, 2, 704–716.
Fabiani, METRO., & Donchin, mi. (1995). Encoding processes and
memory organization: A model of the von Restorff effect.
Revista de Psicología Experimental: Aprendiendo, Memoria,
and Cognition, 21, 224–240.
the response space’’—An account of left prefrontal
activation at encoding. Neuroimagen, 12, 404–417.
Fries, PAG., Reynolds, j. h., Rorie, A. MI., & Desimone, R. (2001).
Modulation of oscillatory neuronal synchronization by
selective visual attention. Ciencia, 291, 1560–1563.
Friston, k. (2003). Learning and inference in the brain.
Neural Networks, 16, 1325–1352.
Gardiner, j. METRO., & Parkin, A. j. (1990). Attention and
recollective experience in recognition memory. Memoria
& Cognición, 18, 579–583.
Gevins, A., & Herrero, METRO. mi. (2000). Neurophysiological
measures of working memory and individual differences
in cognitive ability and cognitive style. Corteza cerebral,
10, 829–839.
Givens, B. S., & Olton, D. S. (1990). Cholinergic and GABAergic
modulation of medial septal area: Effect on working
memory. Neurociencia conductual, 104, 849–855.
Glanzer, METRO., & Cunitz, A. R. (1966). Two storage mechanisms
in free recall. Journal of Verbal Learning and Verbal
Comportamiento, 5, 351–360.
Bruto, J., Schmitz, F., Schnitzler, I., Kessler, K., Shapiro, K.,
Hommel, B., & Schnitzler, A. (2004). Modulation of
long-range neural synchrony reflects temporal limitations
of visual attention in humans. Actas del Nacional
Academia de Ciencias, EE.UU., 101, 13050–13055.
Gruber, T., Muller, METRO. METRO., Keil, A., & Elbert, t. (1999).
Selective visual–spatial attention alters induced gamma
band responses in the human EEG. Clinical
Neurofisiología, 110, 2074–2085.
Gruber, T., Tsivilis, D., Montaldi, D., & Muller, METRO. METRO.
(2004). Induced gamma band responses: An early marker
of memory encoding and retrieval. NeuroReport, 15,
1837–1841.
Hald, l. A., Bastiaansen, METRO. C., & Hagoort, PAG. (2005). EEG
theta and gamma responses to semantic violations in
online sentence processing. Brain and Language.
Herrmann, C. S., Munk, METRO. h., & ángel, A. k. (2004). Cognitivo
functions of gamma-band activity: Memory match and
utilization. Tendencias en Ciencias Cognitivas, 8, 347–355.
Hockley, W.. MI., & Consoli, A. (1999). Familiarity and
recollection in item and associative recognition. Memoria
& Cognición, 27, 657–664.
Huerta, PAG. T., & Lisman, j. mi. (1993). Heightened synaptic
plasticity of hippocampal CA1 neurons during a
cholinergically induced rhythmic state. Naturaleza, 364,
723–725.
Iidaka, T., anderson, norte. D., Kapur, S., Cabeza, r., & Craik,
F. I. (2000). The effect of divided attention on encoding
and retrieval in episodic memory revealed by positron
emission tomography. Revista de neurociencia cognitiva,
12, 267–280.
1130
Revista de neurociencia cognitiva
Volumen 18, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
Kaiser, J., & Lutzenberger, W.. (2005). Human gamma-band
actividad: A window to cognitive processing. NeuroReport,
16, 207–211.
Kensinger, mi. A., Clarke, R. J., & Corkin, S. (2003). Qué
neural correlates underlie successful encoding and retrieval?
A functional magnetic resonance imaging study using a
divided attention paradigm. Revista de neurociencia, 23,
2407–2415.
Klimesch, w., Doppelmayr, METRO., Russegger, h., & Pachinger, t.
(1996). Theta band power in the human scalp EEG and
the encoding of new information. NeuroReport, 7,
1235–1240.
macdonald, A. w., III, cohen, j. D., Stenger, V. A., & Carretero,
C. S. (2000). Dissociating the role of the dorsolateral
prefrontal and anterior cingulate cortex in cognitive
control. Ciencia, 288, 1835–1838.
Makeig, S., Delorme, A., Westerfield, METRO., Jung, t. PAG.,
Townsend, J., Courchesne, MI., & Sejnowski, t. j. (2004).
Electroencephalographic brain dynamics following manually
responded visual targets. Biología PLoS, 2, e176.
Makeig, S., Westerfield, METRO., Jung, t. PAG., Enghoff, S.,
Townsend, J., Courchesne, MI., & Sejnowski, t. j. (2002).
Dynamic brain sources of visual evoked responses.
Ciencia, 295, 690–694.
Mangels, j. A., Picton, t. w., & Craik, F. I. (2001). Atención
and successful episodic encoding: An event-related
potential study. Brain Research, Cognitive Brain Research,
11, 77–95.
McCarthy, GRAMO., & Donchin, mi. (1981). A metric for thought:
A comparison of P300 latency and reaction time. Ciencia,
211, 77–80.
Mesulam, METRO. METRO. (1998). From sensation to cognition. Cerebro,
121, 1013–1052.
Miltner, W.. h., Braun, C., arnold, METRO., Witte, h., & Taub, mi.
(1999). Coherence of gamma-band EEG activity as a basis
for associative learning. Naturaleza, 397, 434–436.
Muller, METRO. METRO., Gruber, T., & Keil, A. (2000). Modulation
of induced gamma band activity in the human EEG
by attention and visual information processing.
International Journal of Psychophysiology, 38,
283–299.
Naveh-Benjamin, METRO., Craik, F. I., Gavrilescu, D., & anderson,
norte. D. (2000). Asymmetry between encoding and retrieval
procesos: Evidence from divided attention and a calibration
análisis. Memoria & Cognición, 28, 965–976.
Naveh-Benjamin, METRO., Guez, J., & Marom, METRO. (2003). El
effects of divided attention at encoding on item and
associative memory. Memoria & Cognición, 31,
1021–1035.
Núñez, PAG. l., Srinivasan, r., Westdorp, A. F., Wijesinghe, R. S.,
Tucker, D. METRO., Silberstein, R. B., & Cadusch, PAG. j. (1997).
EEG coherency. I: Estadísticas, reference electrode, volumen
conduction, Laplacians, cortical imaging, and interpretation
at multiple scales. Electroencephalography and Clinical
Neurofisiología, 103, 499–515.
Otten, l. J., & Donchin, mi. (2000). Relationship between
P300 amplitude and subsequent recall for distinctive
events: Dependence on type of distinctiveness attribute.
Psychophysiology, 37, 644–661.
Otten, l. J., Henson, R. NORTE., & Rugg, METRO. D. (2002). State-related
and item-related neural correlates of successful memory
encoding. Neurociencia de la naturaleza, 5, 1339–1344.
Paller, k. A., Kutas, METRO., & Mayes, A. R. (1987). Neural
correlates of encoding in an incidental learning paradigm.
Electroencephalography and Clinical Neurophysiology,
67, 360–371.
Pfurtscheller, GRAMO. (1992). Event-related synchronization (ERS):
An electrophysiological correlate of cortical areas at rest.
Electroencephalography and Clinical Neurophysiology,
83, 62–69.
posner, METRO. I., & Petersen, S. mi. (1990). The attention system
of the human brain. Revisión anual de neurociencia, 13,
25–42.
Raghavachari, S., Kahana, METRO. J., Rizzuto, D. S., Caplan, j. B.,
Kirschen, METRO. PAG., Bourgeois, B., Madsen, j. r., & Lisman,
j. mi. (2001). Gating of human theta oscillations by a
tarea de memoria de trabajo. Revista de neurociencia, 21,
3175–3183.
Ranganath, C., Yonelinas, A. PAG., cohen, METRO. X., Dy, C. J., Tom,
S. METRO., & D'Esposito, METRO. (2004). Dissociable correlates of
recollection and familiarity within the medial temporal
lobes. Neuropsicología, 42, 2–13.
Rizzuto, D. S., Madsen, j. r., Bromfield, mi. B., Schulze-Bonhage,
A., Seelig, D., Aschenbrenner-Scheibe, r., & Kahana, METRO. j.
(2003). Reset of human neocortical oscillations during a
tarea de memoria de trabajo. Actas del Nacional
Academia de Ciencias, EE.UU., 100, 7931–7936.
Rugg, METRO. D., Otten, l. J., & Henson, R. norte. (2002). El
neural basis of episodic memory: Evidence from functional
neuroimaging. Philosophical Transactions of the Royal
Society of London, Serie B, Ciencias Biologicas, 357,
1097–1110.
Sanquist, t. F., Rohrbaugh, j. w., Syndulko, K., & Lindsley,
D. B. (1980). Electrocortical signs of levels of processing:
Perceptual analysis and recognition memory.
Psychophysiology, 17, 568–576.
Sarnthein, J., Petsche, h., Rappelsberger, PAG., Shaw, GRAMO. l., &
von Stein, A. (1998). Synchronization between prefrontal
and posterior association cortex during human working
memory. procedimientos de la Academia Nacional de Ciencias,
EE.UU., 95, 7092–7096.
Scoville, W.. B., & Milner, B. (1957). Loss of recent memory after
bilateral hippocampal lesions. Journal of Neurochemistry,
20, 11–21.
Montaña de cedro, PAG. B., Kahana, METRO. J., Howard, METRO. w., Donner, mi. J.,
& Madsen, j. R. (2003). Theta and gamma oscillations
during encoding predict subsequent recall. Diario de
Neurociencia, 23, 10809–10814.
Cantante, w., & Gray, C. METRO. (1995). Visual feature integration
and the temporal correlation hypothesis. Annual Review
of Neuroscience, 18, 555–586.
Strange, B. A., Otten, l. J., Josephs, o., Rugg, METRO. D., & Dolan,
R. j. (2002). Dissociable human perirhinal, hippocampal,
and parahippocampal roles during verbal encoding.
Revista de neurociencia, 22, 523–528.
Suckling, J., & bullmore, mi. (2004). Permutation tests for
factorially designed neuroimaging experiments. Humano
Mapeo cerebral, 22, 193–205.
Summerfield, C., & Mangels, j. A. (2005a). Coherent
theta-band EEG activity predicts item-context binding
during encoding. Neuroimagen, 24, 692–703.
Summerfield, C., & Mangels, j. A. (2005b). Functional coupling
between frontal and parietal lobes during recognition
memory. NeuroReport, 16, 117–122.
Tallon-Baudry, C., Bertrand, o., Delpuech, C., & Permier, j.
(1997). Oscillatory gamma-band (30–70 Hz) actividad
induced by a visual search task in humans. Diario de
Neurociencia, 17, 722–734.
Tallon-Baudry, C., Bertrand, o., & pescador, C. (2001).
Oscillatory synchrony between human extrastriate areas
during visual short-term memory maintenance. Diario
of Neuroscience, 21, RC177.
Thompson-Schill, S. l. (2003). Neuroimaging studies of
semantic memory: Inferring ‘‘how’’ from ‘‘where.’’
Neuropsicología, 41, 280–292.
Thompson-Schill, S. l., Bedny, METRO., & Goldberg, R. F. (2005).
Summerfield and Mangels
1131
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
The frontal lobes and the regulation of mental activity.
Opinión actual en neurobiología, 15, 219–224.
Torrence, C., & Campo, GRAMO. PAG. (1998). A practical guide to
wavelet analysis. Bulletin of the American Meteorological
Sociedad, 79, 61–78.
von Restorff, h. (1933). Uber die wirkung von bereichsbildungen
im spurenfeld. Psychologische Forschung, 18, 299–342.
Wagner, A. D., Schacter, D. l., Rotte, METRO., Koutstaal, w.,
Maril, A., Valle, A. METRO., rosa, B. r., & Buckner, R. l.
(1998). Building memories: Remembering and forgetting
of verbal experiences as predicted by brain activity.
Ciencia, 281, 1188–1191.
Weiss, S., & Rappelsberger, PAG. (2000). Long-range EEG
synchronization during word encoding correlates with
successful memory performance. Brain Research,
Cognitive Brain Research, 9, 299–312.
Wickens, C., Kramer, A., Vanasse, l., & Donchin, mi. (1983).
Performance of concurrent tasks: A psychophysiological
analysis of the reciprocity of information-processing
resources. Ciencia, 221, 1080–1082.
Yeung, NORTE., Hombre rico, r., Holroyd, C. B., & cohen, j. D.
(2004). Detection of synchronized oscillations in the
electroencephalogram: An evaluation of methods.
Psychophysiology, 41, 822–832.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
8
mi
7
–
1
pag
1
d
2
F
0
/
1
1
8
9
/
3
5
7
8
/
9
1
4
1
2
oh
0
C
/
norte
1
2
7
0
5
0
6
6
2
1
5
8
5
/
7
j
1
oh
1
C
2
norte
0
.
pag
2
d
0
0
b
6
y
.
gramo
1
tu
8
mi
.
s
7
t
.
oh
1
norte
1
0
2
7
0
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
1132
Revista de neurociencia cognitiva
Volumen 18, Número 7