Interacciones diádicas auditivas a través del “ojo” del cerebro social:
How visual is the posterior STS interaction region?
Julia Landsiedel, Kami Koldewyn
Department of Psychology, School of Human and Behavioural Sciences, Bangor University, Bangor, Reino Unido
Autor correspondiente: Kami Koldewyn (k.koldewyn@bangor.ac.uk)
ABSTRACTO
Human interactions contain potent social cues that meet not only the eye but also the ear. Although research has
identified a region in the posterior superior temporal sulcus as being particularly sensitive to visually presented social
interactions (SI-pSTS), its response to auditory interactions has not been tested. Aquí, we used fMRI to explore brain
response to auditory interactions, with a focus on temporal regions known to be important in auditory processing and
social interaction perception. In Experiment 1, monolingual participants listened to two-speaker conversations (intact
or sentence-scrambled) and one-speaker narrations in both a known and an unknown language. Speaker number
and conversational coherence were explored in separately localised regions-of-interest (ROI). In Experiment 2, bilin-
gual participants were scanned to explore the role of language comprehension. Combining univariate and multivariate
analiza, we found initial evidence for a heteromodal response to social interactions in SI-pSTS. Específicamente, bien
SI-pSTS preferred auditory interactions over control stimuli and represented information about both speaker number
and interactive coherence. Bilateral temporal voice areas (TVA) showed a similar, but less specific, profile. Exploratory
analyses identified another auditory-interaction sensitive area in anterior STS. En efecto, direct comparison suggests
modality specific tuning, with SI-pSTS preferring visual information while aSTS prefers auditory information. Alto-
juntos, these results suggest that right SI-pSTS is a heteromodal region that represents information about social
interactions in both visual and auditory domains. Future work is needed to clarify the roles of TVA and aSTS in audi-
tory interaction perception and further probe right SI-pSTS interaction-selectivity using non-semantic prosodic cues.
Palabras clave: superior temporal sulcus, social interaction, heteromodal, neuroimaging, conversation, narration, MVPA
1.
INTRODUCCIÓN
Every day, humans need to navigate social encounters
which contain potent social cues that meet not only the
ojo (p.ej., gestures and facial expressions) but also the
ear (p.ej., intonation and vocalisation timing). As such,
gauging information from both visual and auditory social
interactions is critical to support adaptive behaviour in a
complex social world. En efecto, human sensitivity to inter-
actions is such that the presence of an interaction facili-
tates processing speed (Papeo et al., 2019; Vestner et al.,
2019), recognition accuracy (Papeo & Abassi, 2019;
Papeo et al., 2017), and working memory efficiency (Ding
et al., 2017; Vestner et al., 2019). Correspondingly, akin to
evidence for face- (Kanwisher & Yovel, 2006), body-
(Downing et al., 2001) or voice-selective (Belin et al.,
2000, 2002) brain areas, neuroimaging studies have iden-
tified interaction-sensitive regions within the bilateral lat-
eral occipito-temporal cortex. Específicamente, the posterior
superior
interaction region
(SI-pSTS) has been found to play a key role in the repre-
sentation of visually perceived dynamic social interac-
ciones. Across a range of stimuli that vary in the strength of
temporal sulcus social
Recibió: 16 Puede 2023 Aceptado: 17 Puede 2023 Available Online: 19 Julio 2023
Neurociencia de la imagen, Volumen 1, 2023
https://doi.org/10.1162/imag_a_00003
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 Septiembre 2023
© 2023 Instituto de Tecnología de Massachusetts. Publicado bajo una atribución Creative Commons 4.0 Internacional (CC POR 4.0) licencia. Research ArticleJ. Landsiedel and K. Koldewyn
Neurociencia de la imagen, Volumen 1, 2023
relevant social cues (p.ej., point-light displays, animated
shapes, videos of dyads), the SI-pSTS responds about
twice as strongly to interacting dyads compared to two
independently acting individuals (Isik et al., 2017; Walbrin
& Koldewyn, 2019; Walbrin et al., 2018) and shows this
selectivity even in naturalistic videos (Landsiedel et al.,
2022; Masson & Isik, 2021). Más, multivariate decod-
ing analyses have found that the right SI-pSTS not only
discriminates between interactors and non-interactors,
but also appears to represent the type and emotional
content of interactions (p.ej., competing/cooperating; Isik
et al., 2017; Walbrin & Koldewyn, 2019; Walbrin et al.,
2018). On the other hand, extrastriate body area (EBA)
has been implicated in the processing of the relational
propiedades; es decir., the facing direction, between two bodies,
which could be classed as “prototypical” visual interac-
ciones (Papeo, 2020), for both static (Abassi & Papeo,
2020, 2021) and dynamic (Bellot et al., 2021) estímulos. Alto-
juntos, this suggests a special role of visually presented
interactive cues within the social brain. Sin embargo, a pesar de
interactions in the world are conveyed through auditory
as well as visual means, auditory cues to interaction have
not received much attention thus far. The current study
sought to address this.
Undeniably, perceiving interactions between others is
not only a visual but also an auditory perceptual experi-
ence. Por ejemplo, a person might overhear two people
behind them conversing with each other or listen to a
radio interview. Even without visual information, a great
deal can be derived about interactions based on not only
what is heard, es decir., the semantic content, but also how
that content is conveyed, es decir., the tone of interaction due
to variations in prosody (p.ej., changes in intonation or
volumen, use of pauses, etc.). En efecto, conversational
characteristics can affect how interactors are perceived
and which characteristics are attributed to them during
conversaciones. For instance, when listening to a two-
speaker conversation which culminates in one person
asking for something, the listener’s ratings of the respon-
dent’s willingness to agree to this request decreased as
the gap between the request and their affirmative
response increased (Roberts & Francisco, 2013; Roberts
et al., 2006). Henetz (2017) used similar procedures and
found that the perception of a conversation’s awkward-
ness as well as interlocutors’ rapport and desire to inter-
act in the future depended on inter-turn silences. Este
emphasises that cues derived whilst listening to interac-
tions are no less informative than cues gathered from
visually observing interactions. De hecho, one could suggest
that most visual cues to interaction have auditory coun-
terpoints that convey nearly identical social information,
including things like the identity of interactants (Awwad
Shiekh Hasan et al., 2016; Stevenage et al., 2012), estafa-
versational turn-taking (Cañigueral & hamilton, 2019;
Pijper & Sanderman, 1994), as well as cues to the inter-
actants’ emotions (de Gelder et al., 2015; Demenescu
et al., 2014; Schirmer & Adolphs, 2017), intentions (Enrici
et al., 2011; Hellbernd & Sammler, 2016), and social traits
(Ponsot et al., 2018; Todorov et al., 2015).
Despite the richness of information contained in audi-
tory interactions, the neural underpinnings of social inter-
investigated almost
action perception have been
exclusively in the visual domain. Studies probing neural
representation of purely auditory interactions are next to
non-existent. The closest proxy are studies investigating
the auditory motion of two people walking, which convey
some sense of togetherness or interactiveness. Bidet-
Caulet et al. (2005) asked participants to listen to foot-
steps of two people walking, one on their left side (izquierda
ear) and one on their right side (right ear). Después,
one of the walkers would cross the auditory scene; de este modo,
their footstep sounds would move towards the same side
as the other walker’s, which required the participants’
respuesta. Compared to a simple noise detection task
(requiring auditory attention), bilateral posterior superior
temporal sulcus (pSTS) increased activation in the foot-
step condition. Sin embargo, their study did not probe audi-
tory interaction perception, per se, which would have
required contrasting the auditory motion of one person vs
two. Work by Saarela and Hari (2008) tested this directly
and found no differences in brain activation in pSTS, o
indeed any other brain region, when comparing footsteps
of two walkers vs one. While these auditory motion stud-
ies do not provide evidence for a region that is selectively
engaged by auditory interactions, to the best of our knowl-
borde, no study has specifically investigated the percep-
tion of auditory interactions using actual conversational
compared to non-conversational speech, or indeed
probed whether regions characterised by its sensitivity to
visual interactions might also be driven by auditory inter-
comportamiento.
In spite of the lack of interaction-specific studies,
investigations focussed on the processing of other social
estímulos (p.ej., faces and voices) support the notion of het-
eromodal (es decir., responding to both visual and auditory
estímulos) processing in the broader STS region (Deen et al.,
2020; Watson et al., 2014), which is in line with reports of
a significant overlap between face- and voice-sensitive
voxels in parts of the pSTS (Deen et al., 2015). Más-
más, several studies have proposed the STS as an area
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 Septiembre 2023
2
j. Landsiedel and K. Koldewyn
Neurociencia de la imagen, Volumen 1, 2023
of audio-visual integration of both emotional and neutral
facial and vocal expressions (Kreifelts et al., 2009; Robins
et al., 2009; Watson et al., 2014; Wright et al., 2003),
reflected by enhanced pSTS activation in response to
both modalities compared to unimodal stimulus presen-
tation. Given its proximity to auditory cortical areas, y
nearby regions demonstrated to be integrative and/or
heteromodal, the SI-pSTS seems an obvious candidate
to investigate in the context of auditory interactions. Este
is in contrast to EBA, which may also be involved in inter-
action processing (Abassi & Papeo, 2020, 2021; Bellot
et al., 2021), but which is considered to be a strictly visual
region and not responsive to auditory information (Beer
et al., 2013).
Across two experiments, we addressed the hypothesis
that the SI-pSTS region might play a crucial role in the pro-
cessing of not only visual but also auditory interactions
using speech stimuli with three levels of interactiveness:
interactions (conversations between two people) and their
scrambled counterparts, as well as non-interactions (sto-
ries narrated by one person) in two languages. En tono rimbombante,
scrambling recombined complete utterances taken from
different interactions. De este modo, speech was comprehensible
at the sentence level but sentences were not semantically
related to each other. In Experiment 1, participants were
monolingual, whereas in Experiment 2, participants were
bilingual. We used functional localisers (Fedorenko et al.,
2010) to define bilateral regions of interest (ROI): el
visual SI-pSTS region, voice-selective temporal voice
areas (TVA), and temporal parietal junction (TPJ) y
tested their responses to auditory interactions across both
univariate and multivariate pattern analyses. We hypothe-
sised that if visual SI-pSTS was, En realidad, heteromodal, él
would show greater response to conversation stimuli
involving two speakers compared to one-speaker narra-
ciones (regardless of semantic comprehension in monolin-
guals in Experiment 1). Beyond this broad test of auditory
interaction sensitivity, we also expected SI-pSTS to be
sensitive to the difference between conversations and
scrambled conversations (which for monolingual partici-
pants deteriorated interactive cues of conversation coher-
ence in their native language, and conversational flow/
prosody in the unknown language). TVA (Agus et al., 2017;
Belin et al., 2000, 2002) was included as an auditory con-
trol region that we expected to respond to all conditions,
though given the dearth of information on auditory interac-
tion processing, we did not have strong expectations
regarding its interaction sensitivity. Our reasoning was that
if SI-pSTS did not show heteromodal characteristics and
sensitivity to auditory interactions could be “found” any-
where in the brain, such sensitivity might emerge in an area
tuned to voices, like TVA. Finalmente, TPJ was included as a
“social” control region that is spatially very near SI-pSTS
but that we did not expect to be driven by either auditory
stimuli in general, or manipulations of interactiveness spe-
cifically (Walbrin & Koldewyn, 2019; Walbrin et al., 2020).
Además, whole-brain analyses were conducted to
explore the wider brain networks implicated in auditory
interaction processing.
2. MÉTODOS
2.1. Experimento 1
2.1.1. Participantes
Twenty-four right-handed participants were recruited to
take part in this study. Handedness was confirmed using
the Edinburgh Handedness Inventory (EHI; Oldfield, 1971).
All participants had normal or corrected to normal vision,
were native English speakers, and had no German language
habilidades. After data exclusion (ver 2.1.2), one participant was
removed from the analyses (final sample of N = 23; significar
age = 22.35, SD = 3.04; 7 machos). All participants gave
informed consent, were debriefed at the end of the study,
and received monetary renumeration for their time. The pro-
tocol was approved by the School of Psychology’s ethics
committee at Bangor University and was pre-registered on
AsPredicted.org (ID23865) en 23/05/2019.
2.1.2. Diseño & procedimiento
To investigate auditory interaction perception with and
without language comprehension, the main experimental
task consisted of a 2 × 3 repeated-measures fMRI event-
related design. Across two languages (English and
Alemán), auditory interactiveness was manipulated using
three conditions: non-interactive one-speaker narrations,
interactive two-speaker conversations, as well as an
intermediate condition using scrambled conversations
(ver 2.1.3 for details). This condition still contained inter-
active cues (two speakers taking turns); sin embargo, conver-
sational content was not coherent. German stimuli were
used to explore interactive effects independent of stimu-
lus comprehension.
Each run contained 36 ensayos, incluido 24 task trials
(4 per condition) y 12 catch trials (2 per condition). El
order of conditions was pseudo-randomised using cus-
tom MATLAB code to optimise the efficiency of the
diseño. The inter-stimulus interval was jittered (mean jit-
ter 1.5 artículos de segunda clase, range = 0–3 seconds). Participants com-
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 Septiembre 2023
3
j. Landsiedel and K. Koldewyn
Neurociencia de la imagen, Volumen 1, 2023
pleted seven runs (28 task trials and 14 catch trials per
condition across runs, de este modo 252 trials overall), cada
between 5.6–5.8 minutes in length. Due to the variability
in stimulus length (range: 6–11 seconds), each run con-
tained a specific set of stimuli. The order of the runs was
counter-balanced across participants.
Participants were instructed to listen attentively, and an
orthogonal catch-trial detection task was used to maintain
(and confirm) participants’ attention throughout. Catch tri-
als consisted of recordings that were manipulated such
that a single word was repeated, p.ej., “Do you need need
anything from the supermarket?". The occurrence of these
repeated catch words was balanced for all conditions
such that the repeated word occurred equally often in the
primero, segundo, tercero, or fourth sentence of a stimulus. Partic-
ipants had to accurately detect seven out of 12 catch trials
per run for that run to be included in subsequent analyses
(see Supplementary S1 for the behavioural results). Basado
on this criterium, two participants had one, and another
participant three run(s) removed from the analyses. Solo
participants with five or more runs after data exclusion
were included in the final sample.
The task was presented in Psychtoolbox 3.0.14
(Brainard, 1997; Kleiner et al., 2007) using MATLAB 2018.a
(The MathWorks Inc.) running on a Linux Ubuntu 16.04
distribution stimulus computer. Sensimetrics (model S15)
MR-safe in-ear earphones were used for stimulus presen-
tation. Stimulus volume was adjusted to a comfortable
level individually for each participant.
2.1.3. Estímulos
Scripted conversations and narrations were developed in
Inglés, and subsequently translated into German by two
native speakers. All stimuli were recorded specifically for
este estudio (see Supplementary S2 for recording details) por
native English or native German speakers. A large set of
stimuli was recorded, from which the final stimulus set was
selected (see Supplementary S3 and S4.1 for details).
Narrations were recorded for each speaker separately.
Narration content was loosely based on and inspired by
children’s books. Care was taken that narrations remained
descriptive rather than invoking mentalising processes.
Conversations were recorded in pairs (two same-
gender pairs per language condition, one male, uno
femenino) to capture “true” interactions. They consisted of
a short exchange (usually four sentences in total) entre
the two speakers taking turns (Agent A – Agent B – Agent
A – Agent B). All conversations were recorded twice so
that both speakers played both agent roles. Conversa-
tions varied in content, p.ej., asking a friend about their
exam or ordering food in a restaurant.
Scrambled conversations were created from the origi-
nal conversation scripts by randomly combining individ-
ual speaker turns from different conversations into a new
combination, which still consisted of two speakers taking
turns, but where the turns were taken from different con-
versations and, de este modo, were unrelated in semantic mean-
En g. En tono rimbombante, this process could result in a speaker
turn being spoken by a different speaker in the scram-
bled compared to the original conversations whilst the
script, es decir., the semantic content remained the same. Para
este, the original conversations (from each gender pair)
were cut up into their individual speaker turns (two per
agent) using Audacity (The Audacity Team) audio soft-
mercancía. Using custom MATLAB code, turns from four ran-
dom original conversations were selected and
re-combined such that each turn was no longer in its
original position of the conversation. Por ejemplo, el
opening turn of a conversation could only appear in the
2nd, 3rd, or 4th position in a scrambled conversation. Este
method was chosen to disrupt the natural conversational
flow not only through mixing up content but also through
disrupting prosodic and intonational cues. Scrambling
generated equal numbers of stimuli, with either speaker
taking the role of agent A or agent B.
All stimuli were analysed in Praat software (http://www
. praat . org) to assess mean pitch (fundamental frequency,
F0) for each condition (see Supplementary S4.2). Más-
más, the final set of English stimuli was rated on per-
ceived naturalness, valence, and imaginability/mental
imagery, and for the two-speaker conditions also on
interactiveness and perceived closeness between the
two speakers. In brief, conversation and narrations were
closely matched on all non-interactive dimensions. En
contrast, conversations and scrambled conversations
significantly differed across all scales (see Supplemen-
tary S4.3 for rating data and statistics).
2.1.4. Localiser tasks
To define independent regions of interest (ROI), participar-
pants also completed a set of established localiser tasks.
The interaction localiser (Isik et al., 2017; Walbrin et al.,
2018) was used to localise the SI-pSTS region sensitive
to visual social interactions. Across three runs, participar-
pants watched videos of two point-light agents in three
condiciones
interactions, y
non-interaction/ independent actions). Each run con-
tained two 16-second blocks per condition and three
(interacción, scrambled
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 Septiembre 2023
4
j. Landsiedel and K. Koldewyn
Neurociencia de la imagen, Volumen 1, 2023
16-second rest blocks (total run time 144 artículos de segunda clase). Bilat-
eral SI-pSTS was localised using the contrast interactions
> non-interactions. The voice localiser (Pernet et al., 2015)
was used to localise the temporal voice areas (TVAs)
along the anterior-posterior axis of the STS. The TVAs
show sensitivity to human vocal (speech and non-speech)
sounds (Agus et al., 2017; Belin et al., 2000, 2002; Pernet
et al., 2015), and seem particularly involved in processing
of paralinguistic information such as gender (Charest
et al., 2013), identity (Latinus et al., 2013), or emotion
(Ethofer et al., 2009, 2012). Por lo tanto, we do not expect to
find interaction sensitivity in the TVAs. Participants com-
pleted one run, listening to human vocal sounds (speech,
p.ej., palabras, or syllables; and non-speech sounds, p.ej.,
laughs or sighs); and non-vocal sounds (natural sounds
like waves or animals, and man-made object sounds like
cars or alarms) across twenty 8-second blocks respec-
activamente. These condition blocks were interspersed with
twenty 10-second blocks of silence (total run time
10.3-minutos). Bilateral TVA was localised using the con-
trast human vocal sounds > non-vocal sounds. Finalmente, a
third localiser, described by Jacoby et al. (2016), era
used to define temporo-parietal junction as a control
regions within the “social brain” (see Supplementary S5
for details).
2.1.5. MRI parameters, pre-processing, & GLM estimation
Data were collected at the Bangor Imaging Centre using
a Philips Achieva 3-T scanner using a 32-channel head
coil (Philips, Eindhoven, Los países bajos). A T2*-weighted
gradient-echo single-shot EPI pulse sequence (with Soft-
Tone noise reduction, TR = 2000 EM, TE = 30 EM) era
used for all tasks (with slightly different parameters
depending on task for flip angle, FOV, number of slices,
and slice order, ver tabla 1).
Structural images were obtained with the following
parámetros: T1-weighted image acquisition using a gra-
dient echo, multi-shot turbo field echo pulse sequence,
with a five echo average; TR = 12 EM, average TE = 3.4 EM,
en 1.7 ms steps, total acquisition time = 136 artículos de segunda clase, flip
angle = 8°, FOV = 240 × 240, acquisition matrix = 240 × 224
(reconstruction matrix = 240); 128 contiguous axial slices,
acquired voxel size (mm) = 1.0 × 1.07 × 2.0 (reconstructed
voxel size = 1 mm3).
Cª). Pre-processing
Pre-processing and general linear model (GLM) esti-
mation were performed using SPM 12 (fil.ion.ucl.ac.uk/
spm/software/spm12) in MATLAB 2018.a (The Math-
Works
included slice-timing
(event-related main auditory task only), realignment and
re-slicing, co-registration to anatomical image, segmen-
tation, normalisation (normalised MNI space with 2 mm
isotropic voxels), and smoothing. All SPM12 default
pre-processing parameters were used, except for the use
of an initial 3 mm FWHM Gaussian smoothing kernel. Este
smoothing kernel is recommended when using ArtRepair
toolbox (v5b; Mazaika et al., 2005). Específicamente, ArtRepair
was used to detect and repair noisy volumes (volumes
that contained more than 1.3% variation in global inten-
sity or 0.5 mm/TR scan-to-scan-motion). Thirteen sub-
jects required repairs in at least one run. Prior to first-level
modelling, data were smoothed again using a 5 mm
FWHM kernel. Subsequently for each run, event or block
durations and onsets for each experimental condition
were modelled using a boxcar reference vector and con-
volved with a canonical hemodynamic response function
(without time or dispersion derivatives) using a 128-sec-
ond high-pass filter and autoregressive AR(1) modelo.
Head motion was modelled using six nuisance regressors
(translation and rotation). Además, for the main audi-
tory task, catch trials were modelled as a regressor of no
interés. Rest periods were modelled implicitly.
2.1.6. Whole-brain group analyses
For localiser tasks, respective contrasts were modelled at
the group level using one-sample t-tests. For the main
auditory task, the Multivariate and Repeated Measures
(MRM) toolbox (McFarquhar et al., 2016) fue usado. Cada
participant’s baseline contrast images of the six experi-
Mesa 1. Scanning parameters
Dummy
scans
Número de
slices
4
5
36, Gap
0 mm
35, Gap
0 mm
Scan order
Ascending
(foot to
cabeza)
Odd-even
(1 3 2 4)
Interaction
localiser
Audio task
Voice
localiser
Flip
angle
FOV
Voxel sizes acquired/
reconstructed
Matrix
83°
240 × 240 × 112
3 × 3.08 ×
Acquisition 80 × 78
3.5 mm/3 mm
Reconstruction 80
240 × 240 × 105
3 × 3.08 ×
77°
3 mm/3 mm
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 Septiembre 2023
5
j. Landsiedel and K. Koldewyn
Neurociencia de la imagen, Volumen 1, 2023
mental conditions were entered into a 2 (Idioma) × 3
(Condition) repeated-measures ANOVA. Within
este
modelo, F-contrasts were computed for the main effects
and the interaction effect using Wilks’ Lambda as multi-
variate test statistic. All reported F-contrasts were thresh-
olded using an initial cluster-forming threshold of p < .001
uncorrected, followed by permutation tests (5000 itera-
tions) to provide cluster-level FWE correction for multiple
comparisons of pFWE < .05.
2.1.7. ROI creation & percent signal change (PSC) analyses
Functional ROIs were defined following a “group-
constrained” ROI definition approach (for details see
Julian et al., 2012). This approach reduces subjectivity
in how ROI locations are selected and ensures that ROI
selection is not influenced by the data that will be
extracted and analysed from a given ROI. To start,
group-level T/F-maps were used to identify MNI coordi-
nates of bilateral ROIs (see Supplementary S6). These
coordinates formed the centres of initial 8 mm bound-
ing spheres. Subject-specific search spaces were then
defined by running a group-level analysis to determine
a peak coordinate for activation that was used to local-
ise this search space using a leave-one-subject-out
(LOSO) approach, i.e., the group contained all subjects
except the “current” subject whose search space was
being defined. The final subject-specific search space
was defined based on the intersection of the original
8 mm bounding sphere with the group-level T-map of
the LOSO scheme. To create the final subject-specific
ROI (see Fig. S1), the top 100 contiguous voxels (high-
est T-values) within the subject-specific search were
selected for each participant individually (Walbrin et al.,
2020). Thus, while all ROIs were 100 voxels in size, they
differed across participants in their exact placement.
Additionally, a leave-one-run-out (LORO) scheme was
implemented in this step (defining an ROI on all but one
run, extracting PSC from the remaining one in an itera-
tive n-fold partition scheme) in cases where ROI defini-
tion and extraction were based on the same task, i.e.,
when testing the response of the SI-pSTS itself to the
interaction localiser. This procedure could not be
applied when testing the response of the TVA to the
voice localiser conditions, however, as the localiser
consisted of only one run. PSC data were extracted
from ROIs using the Marsbar toolbox (Brett et al., 2002).
For the main auditory task, PSC were analysed in a 2
(Language) × 3 (Condition) repeated-measures ANOVAs
for each ROI respectively. Greenhouse-Geisser correc-
tion for violation of assumptions of sphericity was applied
where necessary. Multiple comparison correction was
implemented based on the number of ROIs tested in a
given contrast; i.e., multiple tests for each contrast were
considered as a “family” of statistical tests that should be
corrected across. Given our four regions of interest (bilat-
eral SI-pSTS and TVA), this resulted in a corrected p-value
of p < .0125 (.05/4) for both contrasts used to test main
and interaction effects. For the interaction localiser, dif-
ferences between conditions were analysed using
paired-sample t-tests, with particular focus on two con-
trasts of interest: interactions vs non-interactions and
scrambled interactions respectively. For the auditory
task, multiple comparison correction was applied based
on the number of ROIs tested in a given contrast (cor-
rected p-value of p < .0125). Furthermore, for selected
ROIs, auditory- and visual-interaction selectivity was cal-
culated as the t-value of differences in activation between
conversations vs narrations across languages for the main
experimental task, and interactions vs non-interactions
for the interaction localiser, and compared using paired-
sample t-tests.
For all paired t-test comparisons, effect sizes repre-
sent Cohen′s d for repeated measures (drm), which rep-
resents the mean difference standardised by the standard
deviation of the difference scores corrected for the cor-
relation between the measurements (Lakens, 2013).
2.1.8. Multivariate pattern analyses (MVPA)
Pattern decoding analysis using an iterative n-folds parti-
tion scheme of LORO was implemented using the CoS-
MoMVPA toolbox (Oosterhof et al., 2016) with a focus on
four contrasts of interest: conversations vs narrations and
conversations vs scrambled conversations for each lan-
guage. For each subject, a binary linear support vector
machine (SVM) classifier was trained on a given ROI’s voxel
patterns for the conditions of interest (i.e., beta values in a
subject's respective top 100-voxels ROI, see section 2.1.7,
averaged across all trials per condition per run) in all but
one run of data—with the “left-out” run of data used to
independently test classification performance. This resulted
in as many folds of cross-validation as a subject had
valid task runs (usually seven). Prior to classification, voxel
patterns were normalised (demeaned) for each run sepa-
rately. Following cross-validation, classification accuracy
was averaged across all n-folds iterations before being
entered into group-level analysis. For each contrast of
interest, average classification accuracy was tested against
chance (50%) using one-tailed one-sample t-tests.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
6
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
To correct for multiple comparisons across four ROIs,
we used a so-called “singleton” neighbourhood, where
each ROI to be corrected for was treated as one feature.
This means that each ROI was only a neighbour of itself.
This neighbourhood was then tested using Monte-Carlo
based clustering statistics. Here, we used Thresh-
old-Free-Cluster-Enhancement (TFCE; Smith & Nichols,
2009) as a clustering statistic with 10.000 iterations of
Monte Carlo simulations (cf. cosmo_montecarlo_cluster_
stat.m). Although traditionally, TFCE is used to test cluster
survival based on iteratively testing the spatial clustering at
different height thresholds to determine how much local
support each feature (voxel) has (using a neighbourhood
composed of many features), the same principle can also
be applied to correct for multiple comparison across ROIs
when conceptualising each ROI as a cluster (cf. cosmo_
singleton_neighborhood.m). Each iteration of Monte Carlo
simulations generated null data based on the sample’s
classification accuracies in each ROI using a sign-based
permutation approach (also implemented in FieldTrip; see
Maris and Oostenveld, 2007). Significance was determined
based on the comparison of “clustering” in the null data
across all iterations compared to “clustering” observed in
the original data. This method yields conservative esti-
mates of significance. We report the resulting one-tailed
Z-scores and p-values where Z-scores greater than 1.65
are indicative of significant above chance classification in
each ROI after correction for multiple comparisons. As this
method of multiple comparison correction is somewhat
opaque, we additionally ran standard one-sample t-tests
whose p-values can then be evaluated against a Bonfer-
roni-corrected p-value threshold. Corrected p-values were
determined as above for the PSC analyses, using the num-
ber of ROIs tested in a given contrast (corrected p-value of
p < .0125 based on four ROIs per contrast 0.05/4).
2.2. Experiment 2
2.2.1. Participants
Twelve German native speakers took part in this study
(mean age = 22.50, SD = 2.28, 3 males). All participants
were right-handed as confirmed by the EHI, and had nor-
mal or corrected to normal vision. The sample’s English
language skills were at a minimum of B2 (upper interme-
diate, Common European Framework of Reference for
Languages), which is the minimum level required by uni-
versities for international first-year students. Participants
gave informed consent and were debriefed and paid at
the end of the study.
2.2.2. Design & procedure
All procedures were the same as in Experiment 1,
although participants only completed the main auditory
experimental task as well as the pSTS interaction local-
iser. Due to technical difficulties, the TVA voice localiser
could not be acquired for this sample.
2.2.3. MRI parameters & pre-processing
Compared to Experiment 1, images were acquired on a
different scanner (Philips Igenia Elition X 3T scanner) with
a 32-channel head coil (Philips, Eindhoven, the Nether-
lands) at the Bangor Imaging Centre. Acquisition parame-
ters for functional runs were the same as in Experiment 1.
The structural sequence was slightly different: for each
participant, a high-resolution anatomical T1-weighted
image acquired using a gradient echo, multi-shot turbo
field echo pulse sequence, with a five-echo average;
TR = 18 ms, average TE = 9.8 ms, in 3.2 ms steps, total
acquisition time = 338 seconds, flip angle = 8°, FOV =
224 × 224, acquisition matrix = 224 × 220 (reconstruction
matrix = 240); 175 contiguous slices, acquired voxel size
(mm) = 1.0 × 1.0 × 2.0 (reconstructed voxel size = 1 mm³).
Pre-processing and GLM estimation were performed
using the same pipeline as in Experiment 1. Due to low
levels of head-motion in this sample, ArtRepair was not
used to repair noisy volumes. Due to human error, the
first functional run of the main auditory task had to be
discarded for two participants.
2.2.4. ROI creation & PSC analyses
For the SI-pSTS, the same group constrained ROI defini-
tion process was used as in Experiment 1, resulting in
subject-specific ROIs consisting of the top 100 contigu-
ous voxels in each hemisphere separately. However, due
to the small sample size in this study, the initial 8 mm
constraining sphere used the same group-level MNI
coordinates as in Experiment 1, rather than using coordi-
nates of our underpowered sample. Due to the missing
voice localiser scan, for bilateral TVA, ROIs consisted of
6 mm spheres, again using centre MNI coordinates from
Experiment 1. This radius was chosen to select a sphere
size that contained a comparable number of voxels as
the SI-pSTS ROI (6 mm sphere = 123 voxels). As before,
PSC data were extracted for the experimental conditions
of the main auditory task using Marsbar toolbox for the
respective ROIs and analysed using 2 × 3 repeated-
measures ANOVAs.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
7
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
3. RESULTS
3.1. Experiment 1
Results of the PSC results (Fig. 1 left panel) and MVPA
analyses (Fig. 1 right panel) will be presented below sep-
arately for the visually defined SI-pSTS and auditorily
defined TVA (refer to Table 2 for ANOVA statistics and
Supplementary S7, Table S7 for condition means).
Results for TPJ as an additional control region within the
“social brain” can be found in Supplementary S6, as it
consistently showed activation at or below baseline.
Bspmview toolbox was used for whole-brain data visual-
isation (DOI: 10.5281/zenodo.168074, see also https://
www . bobspunt . com / software / bspmview/).
3.1.1. How does the visually defined SI-pSTS respond to auditory
interactions?
PSC analyses. In line with our predictions, there was
a main effect of Condition, where both conversations
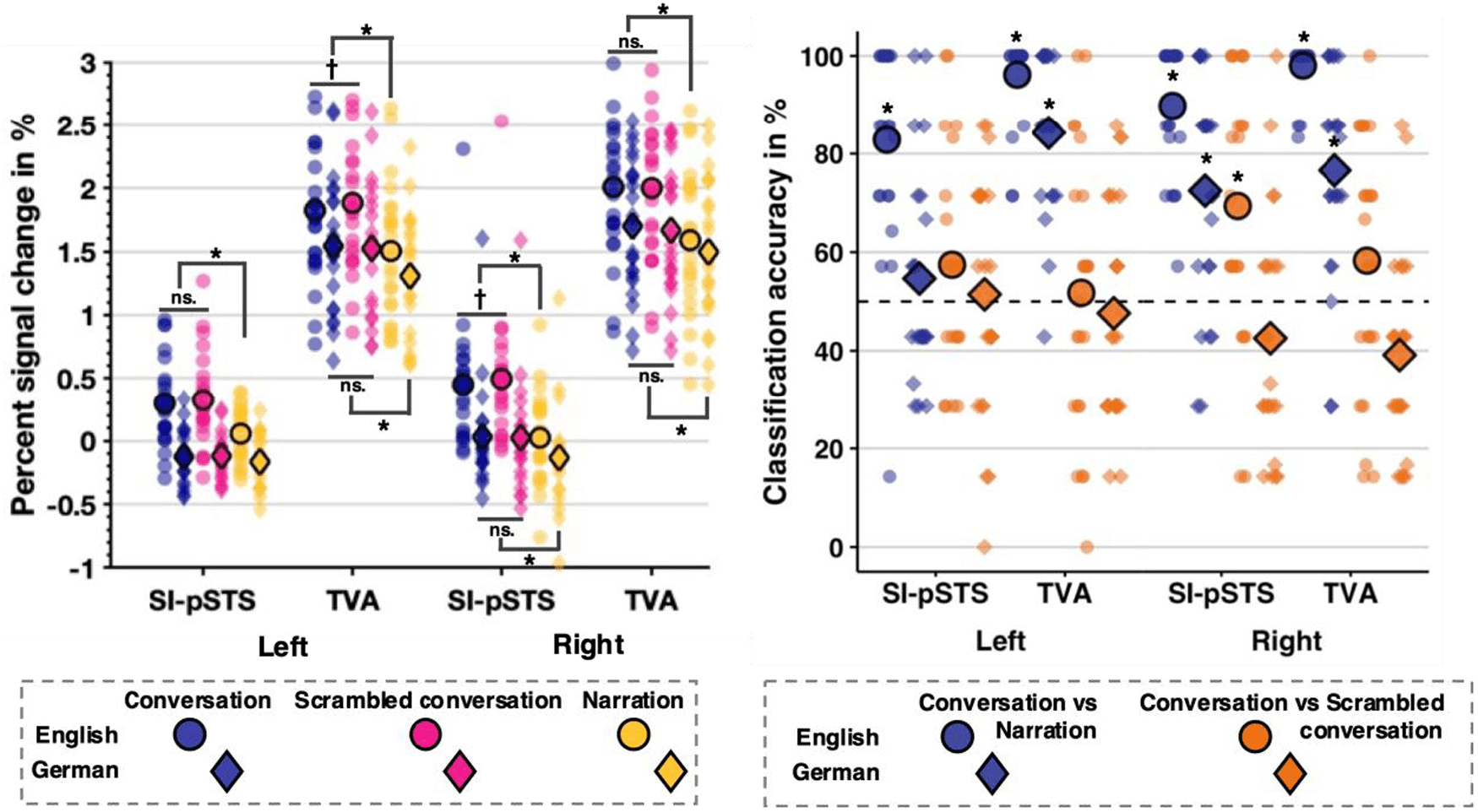
Fig. 1. Condition means (circle/diamond shape with bold edge) and data distribution dot plot for each ROI and hemisphere
for Experiment 1. Left panel: Percent signal change data. Only significant post hoc t-test results are marked (*: p < .0125
corrected, †: p < .05 uncorrected). Effects for English conditions are shown above, and for German conditions below the
condition mean. Right panel: SVM classification accuracy. Significant above chance classification accuracy is indicated using
an asterisk (*: p < .05 TFCE-corrected). Chance level of 50% is represented using the dashed horizontal line.
Table 2. ANOVA results of PSC analyses for the main auditory task in Experiments 1 and 2
Experiment 1
Experiment 2
SI-pSTS
TVA
SI-pSTS
TVA
L
R
L
R
Main effect of language
Main effect of condition
Interaction effect
F
p
2
ηp
F
P
2
ηp
F
p
2
ηp
36.02
<.001
0.62
14.24
<.001
0.39
12.18
<.001
0.36
50.58
<.001
0.70
35.99
<.001
0.62
24.71
<.001
0.53
Italicised p-values indicate significance (p < .05).
15.84
<.001
0.42
40.89
<.001
0.65
25.42
<.001
0.54
29.44
<.001
0.57
62.21
<.001
0.74
9.86
<.001
0.31
8
L
1.37
.27
0.11
10.57
<.001
0.49
0.81
.41
0.07
R
1.71
.22
0.13
23.88
<.001
0.69
3.5
.048
0.24
L
R
4.91
.049
0.31
27.47
<.001
0.71
0.10
.91
0.01
13.56
.004
0.55
20.2
<.001
0.65
0.25
.78
0.02
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
(rSI-pSTS: t(22) = 6.17, p < .001, drm = 0.41, lSI-pSTS:
t(22) = 3.82, p < .001, drm =0.22) and scrambled conversa-
tions (rSI-pSTS: t(22) = 6.13, p < .001, drm = 0.44, lSI-pSTS:
t(22) = 4.00, p < .001, drm = 0.20) evoked greater responses
than narrations in bilateral SI-pSTS. However, PSC did not
differ for intact vs scrambled conversations. Thus, this ef-
fect was driven entirely by a difference between hearing
two speakers vs hearing only one. Unexpectedly, there
was also a large main effect of language, where responses
in bilateral SI-pSTS were greater for English compared to
German stimuli. Indeed, while PSC in bilateral SI-pSTS
was significantly above baseline for English conversations
and English scrambled conversations (all t(22) ≥ 4.06, all
ps < .001), for lSI-pSTS, German conditions led to a signif-
icant decrease in activation (all ts(22) < -2.95, all ps < .007)
and response in rSI-pSTS was not significantly different
than baseline. Both effects suggest that the comprehensi-
bility of heard interactions is important to response within
the SI-pSTS. These main effects were qualified by a signif-
icant Language × Condition interaction. Whilst responses
in lSI-pSTS were only greater for the comprehensible En-
glish two-speaker conversation compared to single-speak-
er narrations (t(22) = 4.14, p < .001, drm = 0.77), rSI-pSTS
showed this pattern independent of comprehensibility (En-
glish: t(22) = 6.28, p < .001, drm = 0.90; German: t(22) = 4.76,
p < .001, drm = 0.41). Additionally, rSI-pSTS response to
English scrambled conversations was slightly greater than
conversations, albeit at an uncorrected
level only
(t(22) = -2.16, p = .04, drm = -0.09), suggesting weak sensi-
tivity to the coherence of comprehensible interactions.
in both
MVPA analyses. Classification analyses in the rSI-pSTS
revealed that the SVM classifier could discriminate
between voxel patterns representing conversations and
(English: M = 0.90,
narrations
languages
SE = 0.03, Z = 3.72, pTFCE < .001, pT-test < .001; German:
M = 0.72, SE = 0.05, Z = 3.24, pTFCE < .001, pT-test < .001),
in line with the PSC results. Crucially, strengthening the
PSC results, the classifier could also decode voxel pat-
terns of English conversations vs scrambled conversa-
tions (M = 0.69, SE = 0.06, Z = 2.49, pTFCE = .006,
pT-test = .002) with above chance accuracy. This suggests
that rSI-pSTS voxel patterns code for interaction infor-
mation based on the number of speakers and interaction
coherence when two speakers are present. Classifica-
tion analyses in lSI-pSTS revealed above chance dis-
crimination between English conversations vs narrations
only (M = 0.83, SE = 0.04, Z = 3.72, pTFCE < .001, pT-test
< .001), suggesting no strong auditory interaction sensi-
tivity in the left hemisphere. German conversations vs
German scrambled conversations were not decodable
above chance in either region.
3.1.2. How does the TVA, a region generally sensitive to voices,
respond to auditory interactions?
PSC analyses. In bilateral TVA, while there was also a sig-
nificant main effect of language, PSC was significantly
greater than baseline for all conditions, regardless of lan-
guage (all t(22) ≥ 12.62, all ps < .001). Thus, bilateral TVA
was clearly driven by voice stimuli regardless of compre-
hensibility. However, bilaterally, PSC was greater in re-
sponse to English compared to German stimuli. As in the
SI-pSTS, there was also a main effect of Condition bilater-
ally. PSC was smaller for narrations compared to both con-
versations (rTVA: t(22) = 7.10, p < .001, drm = 0.54, lTVA:
t(22) = 7.28, p < .001, drm = 0.54) and scrambled conversa-
tions (rTVA: t(22) = 6.09, p < .001, drm = 0.55, lTVA:
t(22) = 9.58, p < .001, drm = 0.51) but no difference was
found between the latter two. Thus, the number of voices,
hearing one or two speakers, clearly modulated TVA acti-
vation. These main effects were qualified by a significant
Language × Condition interaction. Bilaterally, TVA respons-
es were greater for conversations compared to narrations
for both English (rTVA: t(22) = 7.57, p < .001, drm = 0.74,
lTVA:
.001, drm = 0.55) and
German stimuli (rTVA: t(22) = 5.18, p < .001, drm = 0.36,
lTVA: t(22) = 8.98, p < .001, drm = 0.44). Surprisingly, al-
though at an uncorrected level, lTVA responded less to En-
glish conversations compared to scrambled conversations
(t(22) = -2.62, p = .02, drm = -0.09), indicating potential sen-
sitivity to the coherence of comprehensible interactions.
t(22) = 5.66, p <
MVPA analyses. Classification analyses revealed that
voxel patterns representing conversations and narrations
could be decoded for each language respectively in right
(English: M = 0.98, SE = 0.01, Z = 3.72, pTFCE < .001,
pT-test < .001; German: M = 0.77, SE = 0.04, Z = 3.54,
pTFCE < .001, pT-test < .001) and left (English: M = 0.96,
SE = 0.02, Z = 3.72, pTFCE < .001, pT-test < .001; German:
M = 0.84, SE = 0.03, Z = 3.72, pTFCE < .001, pT-test < .001)
TVA (consistent with the PSC results). Importantly, bilater-
ally, discrimination of conversations vs scrambled conver-
sations based on TVA voxel patterns was not successful
for either language.
3.1.3. What is the evidence for heteromodal social interaction
processing in SI-pSTS (and TVA)?
To examine heteromodal processing in response to social
interactions, we examined and, if appropriate, compared
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
9
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
and
scrambled
interactions
responses to visual interactions with responses to auditory
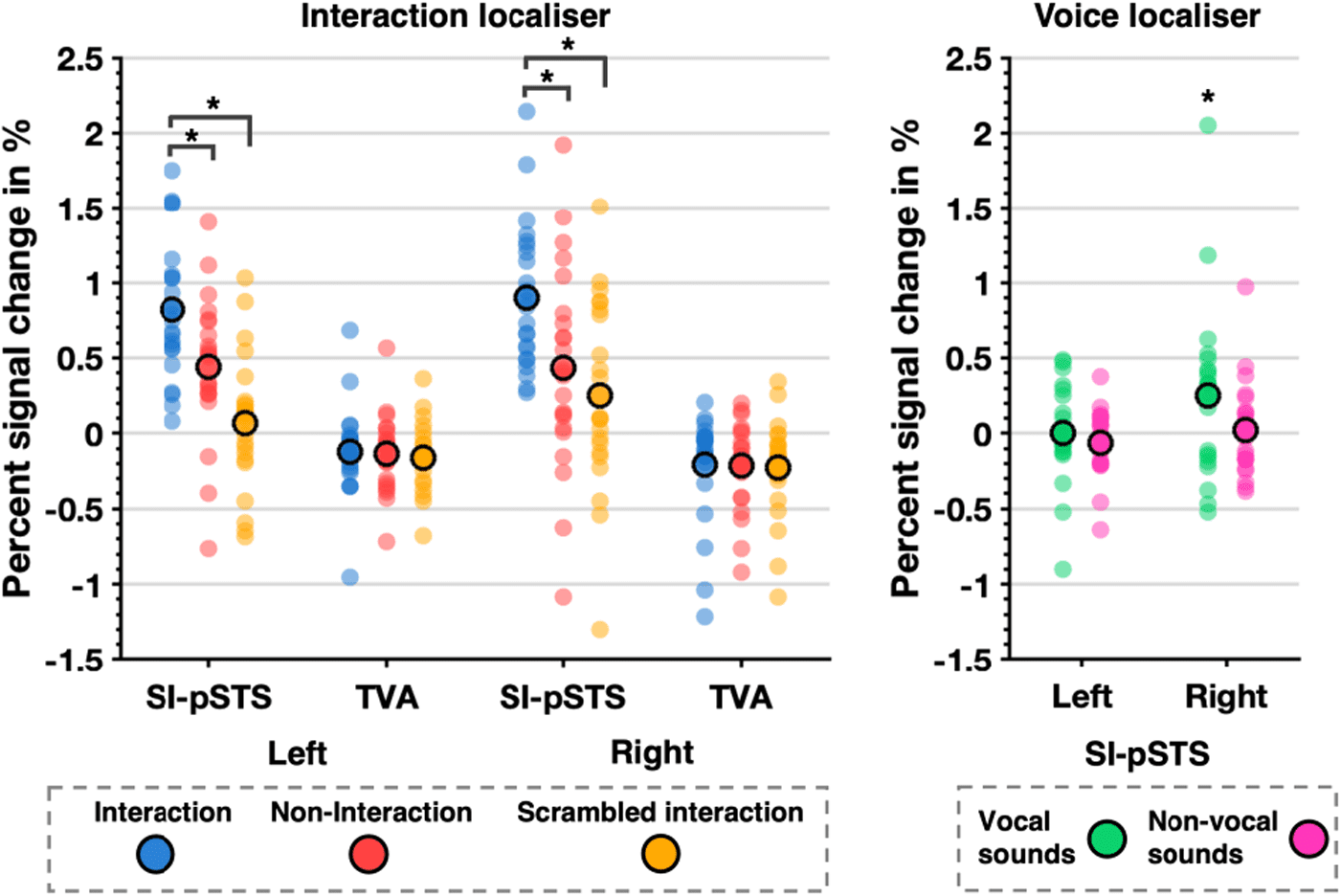
interaction within our ROIs. Analyses confirmed sensitivity
to visual interactions in bilateral SI-pSTS, which showed
greater responses to interactions compared with both
non-interactions
(all
ts(22) > 5.84, all ps < .001; see Fig. 2 left panel and Sup-
plementary S7, Table S8). In contrast, bilateral TVA
responded at or below baseline to the interaction localiser
conditions (all t(23) < -2.00, p < .06). Therefore, the subse-
quent comparison of interaction-selectivity across modal-
ities focussed on SI-pSTS only. Analyses revealed that
visual interaction selectivity (rSI-pSTS: M = 0.62, SE = 0.08,
lSI-pSTS: M = 0.53, SE = 0.06) in bilateral SI-pSTS was
significantly greater (rSI-STS: t(22) = 5.81, p < .001,
drm = 0.34, lSI-pSTS: t(22) = 6.60, p < .001, drm = 0.26) than
auditory interaction selectivity (rSI-pSTS: M = 0.29,
SE = 0.05, lSI-pSTS: M = 0.14, SE = 0.04). Altogether, this
suggests heteromodal processing of social interaction in
SI-pSTS, though with clear preference for visual stimuli.
3.1.4. Does general voice sensitivity explain responses to auditory
interactions in SI-pSTS?
General responsiveness to voice stimuli (see also Sup-
plementary S7, Table S9) was examined by extracting
PSC from the voice localiser in SI-pSTS (see Fig. 2,
right panel). Although vocal sounds activated rSI-pSTS
above baseline (t(23) = 2.16, p = .04), the region was
not strongly driven by human voices. In fact, compared
to comprehensible English conversations (M = 0.45,
SE = 0.10), rSI-pSTS responded about 50% less to
vocal stimuli (M = 0.25, SE = 0.12). Overall, this analysis
suggests that SI-pSTS responses in the main experi-
mental task were not due to a general sensitivity to
voices.
3.1.5. Does whole-brain data reveal an additional region sensitive
to auditory interactions?
To explore whole-brain auditory interaction sensitivity, we
followed a data-driven approach. Rather than focussing
on the main effect of condition, the ROI PSC indicated an
unexpected but robust Language × Condition interaction
effect for our key ROIs. Therefore, the whole-brain interac-
tion effect contrast was used to identify potential candi-
date regions that may be sensitive to auditory interactions.
As it is evident from both Figure 3 and Table 3, brain activ-
ity was modulated by our factors within large clusters in
bilateral STS, including substantial portions of the sulcus
along much of its anterior-posterior axis. Other activations
included prefrontal clusters in bilateral inferior frontal
gyrus, right middle frontal gyrus, left superior medial gyrus,
right anterior cingulate cortex, left precuneus, as well as
the right cerebellum (Crus 2). Bilaterally, the global peak of
F-values fell within the anterior portion of the STS clusters.
In an exploratory post hoc analysis, coordinates close to
Illustration of PSC condition means (circle with bold edge) and data distribution (scatter plot) for localiser data
Fig. 2.
of Experiment 1. Left panel: Interaction localiser data for SI-pSTS and TVA. Significant condition differences are marked
(*p < .0125 corrected). Right panel: Voice localiser data for SI-pSTS only. Above baseline responses are marked (*p < .05).
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
10
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
this peak were used to define bilateral anterior STS (aSTS)
ROIs for PSC extraction using an iterative LORO process
(see 2.1.7 above).
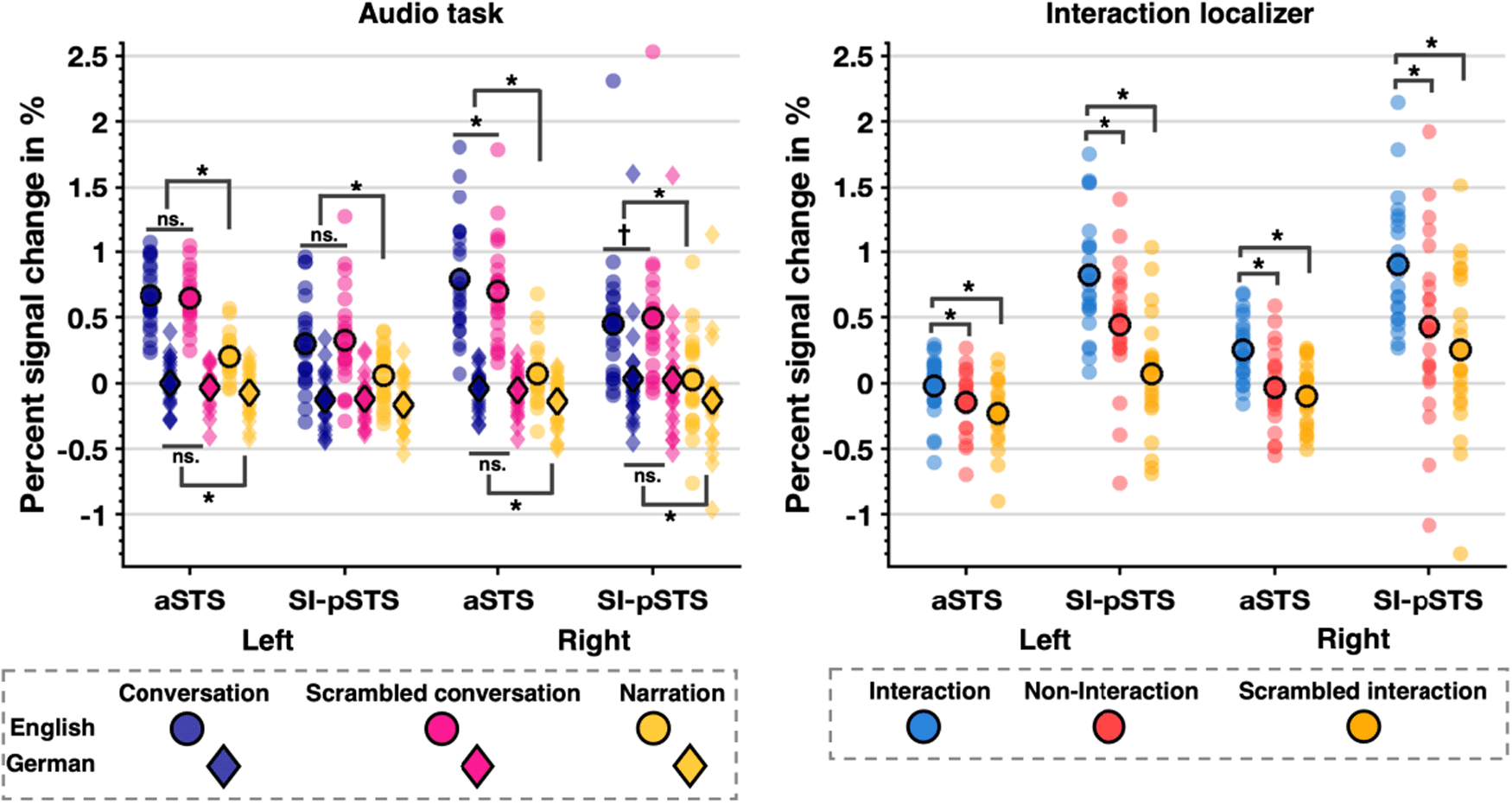
As in both SI-pSTS and TVA regions, PSC analyses
revealed a significant main effect of condition in bilateral
aSTS, indicating sensitivity to auditory interactions (see
Table 4 and Fig. 4 left panel, as well as Supplementary
S7, Table S10 for condition means). PSC was smaller for
narrations (compared to both conversations (raSTS:
t(22) = 9.69, p < .001, drm = 0.22, laSTS: t(22) = 7.29,
p < .001, drm = 0.15) and scrambled conversations (raSTS:
t(22) = 8.60, p < .001, drm = 0.22, laSTS: t(22) = 8.85,
p < .001, drm = 0.13). Interestingly, right but not left aSTS
responded more to conversations compared to scram-
bled conversations (t(22) = 3.08, p = .005, drm = 0.24),
indicating that the region was not merely driven by the
difference of hearing two speakers vs hearing one. Fur-
thermore, there was a main effect of language. Responses
were greater for English compared to German stimuli.
Indeed, PSC in bilateral aSTS was significantly above
baseline for English conversations and English scram-
bled conversations, and for left aSTS also for English nar-
rations (all ts(22) ≥ 6.06, all ps < .001). Additionally,
German narrations led to a significant decrease in activa-
tion (raSTS: t(22) = -3.50, p = .002, laSTS: t(22) = -1.97,
p = .06), whilst for all other conditions, PSC was at base-
line. Thus, aSTS showed a similar effect of comprehensi-
bility as SI-pSTS. Finally, these main effects were qualified
by a significant Language × Condition interaction. For
bilateral aSTS, PSC was greater for conversations
compared to narrations for both English and German
stimuli (all ts(22) > 3.03, p ≤ .006). Finalmente, right aSTS
showed a significantly greater response to conversations
compared to scrambled conversations (t(22) = 2.78,
Higo. 3. Sagittal view of whole-brain group analysis Language × Condition interaction F-contrast. Slices in MNI space with
x-coordinate shown next to each slice.
Mesa 3. Significant clusters for whole-brain language × condition interaction F-contrast, cluster-corrected pFWE < .05
Side
Cluster
Region label
Cluster size
F-value
R
L
1
2
3
4
1
2
3
4
5
6
Medial Temporal Pole
ACC
Cerebellum (Crus 2)
Middle Frontal Gyrus
Middle Temporal Gyrus
Inferior Frontal Gyrus (p. Triangularis)
Precuneus
Superior Medial Gyrus
Posterior Middle Temporal Gyrus
Inferior Frontal Gyrus (p. Triangularis/Opercularis)
2552
102
161
250
1512
147
457
268
463
121
56.54
27.89
26.62
21.84
67.98
29.40
24.88
23.30
19.611
17.844
x
54
16
20
28
-54
-56
-2
-4
-52
-36
y
6
22
-82
36
4
20
-62
46
-62
18
z
-28
26
-36
36
-22
10
26
40
18
24
All x, y, and z coordinates in MNI space.
Table 4. ANOVA results of global aSTS peak ROI PSC analyses for main auditory task in Experiment 1
Main effect of language
Main effect of condition
Interaction effect
Side
F
p
aSTS
L
R
161.20
74.56
<.001
<.001
2
ηp
0.88
0.77
F
p
55.84
78.07
<.001
<.001
2
ηp
0.72
0.78
F
p
54.21
75.66
<.001
<.001
2
ηp
0.71
0.78
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
11
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
Fig. 4. Percent signal change data displaying condition means (circle with bold edge) and data distribution for each
aSTS and SI-pSTS ROI by hemisphere for audio task (left panel) and interaction localiser (right panel). Please note that
pSTS data for the audio task is the same as in Figure 1 and for the interaction localiser is the same as Figure 2. Significant
post hoc t-test results are marked by an asterisk (*: p < .0125 corrected, †: p < .05 uncorrected).
p = .01) for English stimuli only, suggesting sensitivity to
the coherence of comprehensible interactions.
for aSTS (all ts(22) > 3.71, all ps ≤ .001; ver figura. 4 bien
panel and Supplementary S7, Table S11).
3.1.6. What is the evidence for heteromodal social interaction
processing in aSTS?
Further exploratory analyses were conducted to investi-
gate whether a region sensitive to auditory social interac-
tions identified using our main auditory experimental task
would also be responsive to visual social interactions. Como
semejante, this analysis was a reversal of our main experimen-
tal hypothesis, es decir., how does the auditorily defined aSTS
region respond to visual interactions, to explore whether
social interactions are processed cross-modally within
the social brain. De este modo, bilateral aSTS was used to extract
PSC from the interaction localiser.
Visual interactions only activated right aSTS signifi-
cantly above baseline (t(22) = 5.03, pag < .001), whereas
non-interactions (t(22) = -0.61, p = .55) and scrambled
interactions (t(22) = -1.96, p = .06) were at or marginally
below baseline. For left aSTS, all conditions were at
(interactions, t(22) = -0.44, p = .66) or significantly below
(non-interactions t(22) = -2.95, p < .01; scrambled inter-
actions, t(22) = -4.56, p < .001) baseline. Paired-sample
t-tests comparing interactions with non-interactions as
well as interactions with scrambled interactions found
significantly greater responses to interactions bilaterally
Finally, a comparison of interaction-selectivity in the
auditory vs visual domain revealed the reverse pattern to
SI-pSTS (see 3.1.3), greater auditory interaction selectiv-
ity (raSTS: M = 0.83, SE = 0.09, laSTS: M = 0.54,
SE = 0.07) compared to visual interaction selectivity
(raSTS: M = 0.29, SE = 0.05, laSTS: M = 0.12, SE = 0.07)
(raSTS: t(22) = -7.07, p < .001, drm = 0.36, laSTS:
t(22) = -5.94, p < .001, drm = 0.29).
3.1.7. Summary
The main aim of this experiment was to investigate
whether the interaction-sensitive SI-pSTS region is not
only responsive to visual interactions but also responsive
to auditory interactions. Both univariate and multivariate
ROI analyses suggest that bilateral SI-pSTS displays
interaction sensitivity to a broad contrast of two speakers
vs one speaker. Univariate results also lend tentative
support that right SI-pSTS exhibits interaction sensitivity
beyond the number of speakers. This notion was corrob-
orated more strongly using decoding analyses. Specifi-
cally, right SI-pSTS was the only region which could
decode conversations vs scrambled conversations, indi-
cating that it also represents information about the
meaningfulness of an auditory interaction. Unexpectedly,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
12
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
there were strong effects of language: bilateral TVA
responded above baseline across both languages,
whereas SI-pSTS was not driven by German stimuli. In
contrast to our predictions, it seems likely that language
comprehension was an important factor in some of our
results. However, right SI-pSTS could discriminate
between German conversations and narrations, sug-
gesting that comprehension is not a pre-requisite when
processing interactions at the level of speaker number.
Finally, bilateral TVA also exhibited sensitivity to interac-
tion based on the number of speakers, and unexpect-
edly, like SI-pSTS, left TVA also exhibited weak interaction
sensitivity beyond the number of speakers, but only in
univariate analyses.
Furthermore, this experiment explored (1) whether
there was another brain region particularly sensitive to
auditory interactions, and (2) whether visual and/or audi-
tory interaction-sensitive regions may exhibit a hetero-
modal response profile. We used whole-brain group
response to “find” a region in bilateral aSTS and explored
its auditory and visual interaction sensitivity. Interestingly,
right aSTS displayed a response profile characterised by
greater sensitivity to auditory than visual interactions,
whereas the right SI-pSTS showed the opposite pattern
of greater sensitivity to visual compared to auditory inter-
actions. Importantly, both regions showed sensitivity to
interactive content across modality.
3.2. Experiment 2
This experiment was conducted as a small-scale follow-
up study to address the unexpectedly strong language
effects observed in Experiment 1. Here, participants were
fluent in English but not German; thus, language compre-
hension might have driven PSC responses. For instance,
SI-pSTS bilaterally was either at or below baseline for
German conditions. This might be the result of listening
to recordings in a language one does not comprehend in
the context of a language you understand very well within
the same run. A similar native vs unknown language com-
prehension effect has been found in prior work (Cotosck
et al., 2021) using a target word detection task whilst lis-
tening to stories. On the other hand, language-specific
acoustic differences (Mennen et al., 2012, see also Sup-
plementary S4.2) might have driven some of these differ-
ences, particularly in the TVA. Experiment 2 set out to
address this question with particular focus on the SI-pSTS
by using the same stimulus set but testing German-En-
glish bilingual participants.
Please refer to Table 2 for ANOVA statistics, Supple-
mentary S7, Table S7 for condition means, and Figure 5
for an illustration of the PSC results of Experiment 2.
3.2.1. Does SI-pSTS respond differently across languages when
both are understood?
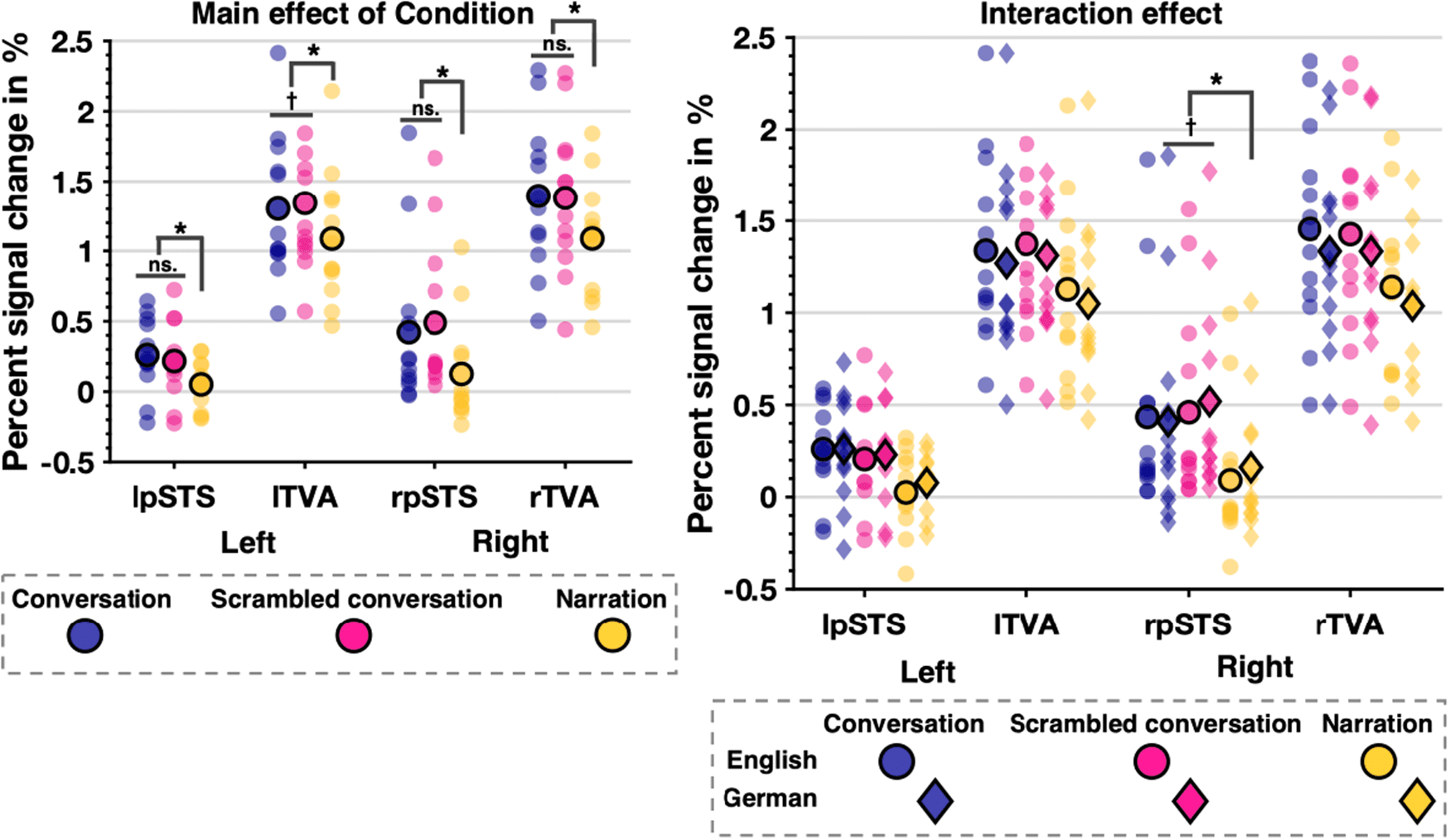
When both languages were comprehensible to partici-
pants, there was no main effect of language. Responses
in bilateral SI-pSTS were similar for English and German
stimuli. Indeed, PSC in bilateral SI-pSTS was significantly
above baseline for both English and German conversa-
tions and scrambled conversations (all t(11) ≥ 2.36, all
ps < .04). Thus, regardless of language, only narrations
did not activate the SI-pSTS. Replicating Experiment 1,
there was a main effect of condition, driven by a differ-
ence between hearing two speakers vs hearing only one.
PSC was smaller for narrations compared to both con-
versations (rSI-pSTS: t(11) = 4.41, p < .001, drm = 1.04,
lSI-pSTS: t(11) = 3.66, p < .001, drm = 0.25) and scram-
bled conversations (rSI-pSTS: t(11) = 6.35, p < .001,
drm = 0.80, lSI-pSTS: t(11) = 3.17, p < .001, drm = 0.27) but
not different between conversations and scrambled con-
versations. These main effects were qualified by a signif-
icant Language × Condition interaction in the right
SI-pSTS only. PSC was greater for conversations com-
pared to narrations for both English (t(11) = 5.23, p < .001,
drm = 0.41) and German stimuli (t(11) = 3.31, p = .007,
drm = 0.26). In contrast, response to German (t(11) = -2.38,
p = .04, drm = 0.18) but not English (t(11) = -0.59, p = .57)
scrambled conversations was significantly greater than
conversations, albeit at an uncorrected level only. This
suggests some sensitivity to the coherence of interac-
tions in the participants’ native language.
3.2.2. How does the TVA response compare in this case?
As in Experiment 1, all conditions strongly activated bilat-
eral TVA above baseline (all t(11) ≥ 7.69, all ps < .001).
Bilaterally, although marginally in the left hemisphere, a
significant effect of language remained even when partic-
ipants understood both languages. Similarly, PSC was
greater for English compared to German conditions. Fur-
ther replicating Experiment 1, there was also a main
effect of condition. PSC was smaller for narrations com-
pared to both conversations (rTVA: t(11) = 4.70, p < .001,
drm = 0.57, lTVA: t(11) = 4.82, p < .001, drm = 0.52) and
scrambled conversations (rTVA: t(11) = 4.53, p < .001,
drm = -0.18, lTVA: t(11) = 5.89, p < .001, drm = 0.56). Finally,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
13
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
Fig. 5. Experiment 2 PSC data illustrating the main effect of Condition (*: p < .017 corrected) and Language × Condition
interaction effect (*: p < .0125 corrected, †: p < .05 uncorrected).
sustaining the unexpected finding from Experiment 1 of
lTVA sensitivity to not only number of speakers but also
coherence of conversation, responses to scrambled con-
versations were slightly but significantly greater than for
intact conversations in left TVA only, albeit at an uncor-
rected level (t(11) = -2.61, p < .03, drm = 0.56). No signifi-
cant interaction effect emerged.
cessing of the participants’ second language (Hasegawa
et al., 2002). Like Experiment 1, left TVA showed a greater
response to scrambled compared to intact conversa-
tions. Thus, across both experiments, left TVA was less
responsive to meaningful conversations.
4. DISCUSSION
3.2.3. Summary
Experiment 2 set out to test whether language compre-
hension may have driven some of the effects seen in
Experiment 1. Testing bilingual participants revealed that
when participants comprehended both languages, lan-
guage effects disappeared in bilateral SI-pSTS whilst
condition effects remained. In line with the results from
Experiment 1, SI-pSTS responded more strongly to con-
versations compared to narrations. Crucially, right
SI-pSTS was sensitive to the difference between German
conversations and German scrambled conversations.
This replicates Experiment 1 which found this difference
for English stimuli. Taken together, these findings suggest
that right SI-pSTS is sensitive to meaningful auditory
interactions, at least in participants’ native language. For
bilateral TVA, language effects remained relatively stable
with the participants’ non-native language resulting in
greater activation. This might point to more effortful pro-
Whilst everyday social
interactions provide a rich
multi-sensory experience, neuroimaging studies of social
interaction perception have predominantly focussed on
the abundance of visual cues they provide. Conversely,
not much is known about auditory interaction perception
in the social brain. Combining univariate and multivariate
analyses, we confirmed our key prediction that visual
SI-pSTS exhibits heteromodal processing of social inter-
actions. In contrast, although voice-selective TVA shows
an unexpectedly similar response profile to auditory
interactions, it is clearly a unimodal region. Specifically,
both bilateral SI-pSTS and TVA were sensitive to interac-
tive information in a broad contrast between two-speaker
conversations and one-speaker narrations, in line with
similar recent work focussed on language processing
(Olson et al., 2023). More importantly, right SI-pSTS
and left TVA showed some weak sensitivity to auditory
interactions when coherence of comprehensible (native
language) conversations was manipulated. However,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
14
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
multivariate decoding analyses only corroborated this
finding for right SI-pSTS, suggesting that the information
represented in SI-pSTS voxel patterns is qualitatively dif-
ferent from that represented in TVA.
These findings are in line with previous results that put
the broader pSTS region at the heart of heteromodal or
even multimodal integrative processing of social informa-
tion (Kreifelts et al., 2009; Lahnakoski et al., 2012; Robins
et al., 2009; Watson et al., 2014; Wright et al., 2003).
Indeed, regions along the pSTS show tuning to a variety
of both visual and auditory social stimuli (Deen et al,
2015) and the pSTS is widely referred to as the “hub” of
the social brain because of its involvement across varied
social tasks (e.g., Yang et al., 2015). However, much of
the prior literature has investigated heteromodal process-
ing in the context of social signals from individuals, mak-
ing this study’s focus on the perception of social
interactions relatively unique. Importantly, our data make
clear that auditory interaction sensitivity in right SI-pSTS
reflects more than tuning to voice stimuli in general.
Indeed, the SI-pSTS region shows negligible sensitivity
to vocal sounds in response to the voice localiser (see
Fig. 2). At the same time, responses in SI-pSTS to visual
and auditory interactive stimuli were not fully equivalent.
Right SI-pSTS interaction selectivity for visual stimuli was
about 50% greater than for auditory stimuli. It could be
that the nature of the interaction-region localiser might, in
part, account for this. Essentially, we tested how SI-pSTS
voxels sensitive to interaction information conveyed by
human body- and biological motion cues responded to
interaction cues conveyed by voice. Body and voice
cues, however, are less strongly associated with each
other compared to face and voice cues. Had we used
stimuli that relied on facial cues of interaction in our local-
iser, we might have found a greater degree of correspon-
dence between visual and auditory SI-pSTS response
profiles. Indeed, heteromodal responses in the broader
STS region to voices have previously been shown in
conjunction with face stimuli (Deen et al., 2015, 2020;
Watson et al., 2014), though not in the context of social
interactions. As such, our approach is a strong test of
whether SI-pSTS shows sensitivity to interactive informa-
tion across modalities.
Nonetheless, the response profile of SI-pSTS to audi-
tory interactions was more nuanced and less definitive
than originally predicted. As a broad test of auditory inter-
action sensitivity, we expected that the mere presence of
two speakers taking conversational turns would drive
SI-pSTS activation regardless of language comprehen-
sion. However, testing monolinguals (Experiment 1) and
bilinguals (Experiment 2) revealed that comprehension
mattered. The SI-pSTS was only driven by the two-speaker
conditions in monolingual participants’ native language,
whereas this language effect was abolished in bilingual
speakers. Nevertheless, in monolingual English speakers,
MVPA analysis of voxel patterns of SI-pSTS revealed that
the two-speaker conditions could be discriminated from
narrations even in the German condition. While this is per-
haps not surprising, it does suggest that language com-
prehension is not a pre-requisite for representation of
information; i.e., number of speakers, that clearly distin-
guishes auditory interactions from non-interactions within
the SI-pSTS. When this distinction is less obvious how-
ever, interactive cues might well be derived through lan-
guage comprehension. Indeed, monolingual participants
listening to intact and scrambled conversations presented
in their native language would be able to differentiate them
based on detecting conversational coherence and pres-
ence of overall gist, whereas without comprehension, they
would have to rely on subtle prosodic cues. We found that
right SI-pSTS only distinguished between the two-speaker
conditions when participants could access their meaning.
Thus, language comprehension clearly mattered when
extracting cues to conversational coherence. Notably,
SI-pSTS lies in proximity to a bilateral brain network
(including STG, STS, MTG, and left IFG, see Bookheimer,
2002; Mar, 2011; Vigneau et al., 2011; Walenski, 2019;
Yang, 2014) implicated in higher-level discourse compre-
hension processes such as evaluation of global coher-
ence, pragmatic interpretations, and text integration at the
gist-level. However, right SI-pSTS’s overlap with this net-
work is unclear. Although domain-general language pro-
cesses designed to detect coherence could contribute to
the response difference between intact and scrambled
interactions, they cannot explain the drop in response to
coherent narrations. Instead, right SI-pSTS might receive
and integrate coherence or gist information from nearby
language regions as a cue to evaluate interactiveness. Fur-
thermore, although we found no support that SI-pSTS was
sensitive to conversational flow in an unknown language,
strong between-language effects may have overshad-
owed the potential to detect more subtle effects of pros-
ody. Future studies investigating the role of SI-pSTS
independent of language comprehension are needed to
firmly establish its role when cues to interaction are harder
to extract; e.g., using low-pass filtered muffled stimuli con-
taining only prosodic but no lexico-semantic cues to inter-
action.
Unexpectedly, voice-selective TVA, especially in the left
hemisphere, exhibited a similar response profile to right
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
15
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
SI-pSTS. Firstly, a greater response to and decoding of
conversations vs narrations across languages, and sec-
ondly, a slightly greater response to scrambled conversa-
tions compared to conversations. Importantly, however,
classification analyses in left TVA could not discriminate
scrambled from intact conversations. Thus, it is unclear
how distinctly (left) TVA responses could be attributed to
pure interaction sensitivity. Indeed, it is possible that
response difference between the one-speaker narration
condition and the two-speaker conditions (conversations,
scrambled) in both TVA and SI-pSTS could partially be
driven by these regions adapting to vocal quality or
speaker identity in the narration condition. However, pSTS
responsivity to voices is thought to reflect higher-level
social process because individuals with lesions in pSTS
are still able to discriminate between and recognise indi-
vidual voices (Jiahui et al, 2017). In addition, we think it
unlikely that adaptation can fully explain our effects in the
SI-pSTS because we do not see strong differences
between these conditions when participants do not under-
stand what is being said, making simple adaptation effects
unlikely. However, as TVA is known to be involved in the
spectro-temporal analysis of human vocal sounds (and
speech) (Agus et al., 2017; Belin et al., 2000, 2002), adap-
tation to vocal quality or identity may partially explain our
effects in this region. Similarly, previous research has
found a significant positive association between mean F0
of speech and TVA activation (Wiethoff et al., 2008). More
generally, TVA is part of the STS/STG engaged in phono-
logical language processing (Vigneau et al., 2006, 2011).
Notably, due to the complexity and diversity of the stimuli
used in this study, acoustic features of the stimulus set
could not be as tightly controlled as we might have liked,
which may drive some between-condition differences in
TVA activation. Specifically, F0 was greater for conversa-
tions compared to narrations, and greater for English com-
pared to German stimuli (see Supplementary S4.2). Thus,
greater TVA activation to conversations compared to nar-
rations, and English compared to German observed across
both experiments could be at least partially explained by
their corresponding differences in F0. However, as intact
and scrambled conversations were matched on F0, these
differences cannot explain higher left TVA response to
scrambled conversations. Importantly, although scram-
bled and intact conversations contained identical sen-
tences, they were not exact phonological equivalents as
our scrambling process allowed a change in speaker.
Thus, differential responses in left TVA might reflect sensi-
tivity to speaker-dependent variations
in phonation
between conditions. Importantly, the above explanations
would not apply to responses in right SI-pSTS, which is
not known to be involved in phonological processes
(Vigneau et al., 2006, 2011) or modulated by F0 (Wiethoff
et al., 2008). In addition, between-language differences
were no longer present in the SI-pSTS when participants
understood both languages (Experiment 2). Altogether, our
findings might motivate future work into auditory interac-
tion perception in the brain to corroborate evidence on the
unique role of right SI-pSTS using more tightly controlled
stimuli that could additionally clarify the role of TVA.
Finally, we confirmed our prediction that TPJ, a social
cognition region selectively engaged by mentalising pro-
cesses (Saxe et al., 2009; Van Overwalle & Baetens, 2009),
is neither driven by our stimuli nor sensitive to differences
between conditions (see Supplementary S5 for details). TPJ
can be engaged by auditory stimuli when participants are
engaged in a mentalising task (Kandylaki et al., 2015; Saxe
et al., 2009). However, while some prior work has suggested
that TPJ is involved in processing social interactions (e.g.,
Canessa et al., 2012; Centelles et al., 2011), it is likely that
the region is involved only when mentalising is required
(Masson & Isik, 2021; Walbrin et al., 2018), which was not
the case in our task. Indeed, we took care to select stimuli
that did not imply nor require mentalising. Similarly, our
whole-brain analysis suggests that occipital and temporal
areas outside the STS, including EBA, are not involved in
auditory interaction perception. This makes sense, as
“early” social perception regions like EBA are not usually
considered to be heteromodal or, indeed, particularly
responsive to auditory stimuli (Beer et al., 2013). Instead,
our exploratory whole-brain analysis identified an area in the
right anterior STS, close to the temporal poles, which
showed sensitivity to interactive information both through
greater activation to two speakers than to one and through
a higher response to comprehensible conversations com-
pared to scrambled conversations (Experiment 1). While
this region’s responsiveness to auditory interactions (and
perhaps to interactive information in general) needs to be
replicated (though see Olson et al, 2023), this is a particu-
larly intriguing finding in light of prior work that highlights the
dorsolateral anterior temporal lobe (ATL) as a region that
may be involved in social semantics (Arioli et al, 2021; Lin
et al., 2018; Zhang et al., 2021; but see also Balgova et al.,
2022; Binney et al., 2016; Binney & Ramsey, 2020 for a
broader perspective regarding the role of the ATL in social
cognition). Furthermore, in the context of speech compre-
hension, it has been suggested that the meaning of speech
is processed in bilateral anterior temporal cortex, including
aSTS (e.g., Mitchell et al., 2003; Scott et al., 2000, 2009,
for a review see Price, 2012). Thus, right aSTS might be
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
16
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
particularly involved in the semantic analysis of auditory
interactions, given that its activation was not driven by com-
prehensible narrative stimuli.
5. CONCLUSION
Our results present initial evidence that SI-pSTS, initially
defined visually, is also sensitive to interactive cues pre-
sented in the auditory domain. In other words, this region
is characterised by a heteromodal response profile that
appears to be particularly sensitive to social interactions.
Future research is needed to both replicate these novel
findings and look beyond the number of speakers and
interaction coherence to investigate whether SI-pSTS
codes for other auditory cues involved in understanding
social interactions, for instance, subtle prosodic cues or
interactional turn duration. In addition, our results may
motivate future work to determine how SI-pSTS inte-
grates multimodal audio-visual social interaction infor-
mation to inform our understanding of highly naturalistic
everyday life social interactions. Finally, this initial work
also prompts further research into the role of aSTS
regions in social interaction perception more broadly and
in conversation/language-based interactions specifically.
DATA AND CODE AVAILABILITY
Stimulus examples as well as subject-level summary data
that support the findings of this study are openly available
at the project’s OSF page at https://osf . io / 4xedj/.
AUTHOR CONTRIBUTIONS
Julia Landsiedel: Conceptualisation, Methodology, Inves-
tigation, Formal analysis, Project administration, Visuali-
sation, Writing—Original Draft, and Writing—Review &
Editing; Kami Koldewyn: Conceptualisation, Funding
acquisition, Methodology,
Investigation, Supervision,
Project administration, Writing—Original Draft, and
Writing—Review & Editing.
FUNDING
This work has received funding from the European
Research Council under the European Union’s Horizon
2020 research and innovation programme (ERC-2016-
STG-716974: Becoming Social).
DECLARATION OF COMPETING INTEREST
None.
ACKNOWLEDGEMENTS
The authors are grateful to Corinne Voigt-Hill for her help
with data collection as well as to members of the Social
Neuroscience and Cognition group and Bangor Imaging
Group at Bangor University for general feedback, helpful
discussion, and suggestions throughout the research
process.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available with
the online version here: https://doi . org / 10 . 1162 / imag _ a
_ 00003.
REFERENCES
Abassi, E., & Papeo, L. (2020). The representation of two-
body shapes in the human visual cortex. J Neurosci,
40(4), 852–863. https://doi . org / 10 . 1523 / JNEUROSCI
. 1378 - 19 . 2019
Abassi, E., & Papeo, L. (2021). A new behavioral and neural
marker of social vision. bioRxiv, 2021.2006.2009.447707.
https://doi . org / 10 . 1101 / 2021 . 06 . 09 . 447707
Agus, T. R., Paquette, S., Suied, C., Pressnitzer, D., & Belin,
P. (2017). Voice selectivity in the temporal voice area
despite matched low-level acoustic cues. Sci Rep, 7(1),
11526. https://doi . org / 10 . 1038 / s41598 - 017 - 11684 - 1
Arioli, M., Gianelli, C., & Canessa, N. (2021). Neural
representation of social concepts: A coordinate-based
meta-analysis of fMRI studies. Brain Imaging Behav,
15(4), 1912–1921. https://doi . org / 10 . 1007 / s11682 - 020
- 00384 - 6
Awwad Shiekh Hasan, B., Valdes-Sosa, M., Gross, J.,
& Belin, P. (2016). “Hearing faces and seeing voices”:
Amodal coding of person identity in the human brain. Sci
Rep, 6, 37494. https://doi . org / 10 . 1038 / srep37494
Balgova, E., Diveica, V., Walbrin, J., & Binney, R. J. (2022).
The role of the ventrolateral anterior temporal lobes in
social cognition. Hum Brain Mapp, 43(15), 4589–4608.
https://doi . org / 10 . 1002 / hbm . 25976
Beer, A., Plank, T., Meyer, G., & Greenlee, M. (2013).
Combined diffusion-weighted and functional magnetic
resonance imaging reveals a temporal-occipital network
involved in auditory-visual object processing. Front Integr
Neurosci, 7, 5. https://doi . org / 10 . 3389 / fnint . 2013 . 00005
Belin, P., Zatorre, R. J., & Ahad, P. (2002). Human temporal-
lobe response to vocal sounds. Brain Res Cogn Brain
Res, 13(1), 17–26. https://doi . org / 10 . 1016 / s0926
- 6410(01)00084 - 2
Belin, P., Zatorre, R. J., Lafaille, P., Ahad, P., & Pike, B.
(2000). Voice-selective areas in human auditory cortex.
Nature, 403(6767), 309–312. https://doi . org / 10 . 1038
/ 35002078
Bellot, E., Abassi, E., & Papeo, L. (2021). Moving toward
versus away from another: How body motion direction
changes the representation of bodies and actions in the
visual cortex. Cereb Cortex, 31(5), 2670–2685. https://doi
. org / 10 . 1093 / cercor / bhaa382
Bidet-Caulet, A., Voisin, J., Bertrand, O., & Fonlupt, P. (2005).
Listening to a walking human activates the temporal
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
17
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
biological motion area. Neuroimage, 28(1), 132–139.
https://doi . org / 10 . 1016 / j . neuroimage . 2005 . 06 . 018
Binney, R. J., Hoffman, P., & Lambon Ralph, M. A. (2016).
Mapping the multiple graded contributions of the anterior
temporal lobe representational hub to abstract and social
concepts: Evidence from distortion-corrected fMRI.
Cereb Cortex, 26(11), 4227–4241. https://doi . org / 10
. 1093 / cercor / bhw260
Binney, R. J., & Ramsey, R. (2020). Social semantics: The
role of conceptual knowledge and cognitive control in
a neurobiological model of the social brain. Neurosci
Biobehav Rev, 112, 28–38. https://doi . org / 10 . 1016 / j
. neubiorev . 2020 . 01 . 030
Bookheimer, S. (2002). Functional MRI of language: New
approaches to understanding the cortical organization of
semantic processing. Annu Rev Neurosci, 25, 151–188.
https://doi . org / 10 . 1146 / annurev . neuro . 25 . 112701 .
142946
Brainard, D. H. (1997). The psychophysics toolbox.
Spat Vis, 10(4), 433–436. https://doi . org / 10 . 1163
/ 156856897X00357
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B.
(2002). Region of interest analysis using an SPM toolbox
8th International Conference on Functional Mapping
of the Human Brain, Sendai, Japan. https://matthew.
dynevor.org/research/abstracts/marsbar/marsbar_
abstract.pdf
Canessa, N., Alemanno, F., Riva, F., Zani, A., Proverbio, A. M.,
Mannara, N., Perani, D., & Cappa, S. F. (2012). The neural
bases of social intention understanding: The role of
interaction goals. PLoS One, 7(7), e42347. https://doi . org
/ 10 . 1371 / journal . pone . 0042347
Cañigueral, R., & Hamilton, A. F. d. C. (2019). The role of
eye gaze during natural social interactions in typical and
autistic people. Front Psychol, 10. https://doi . org / 10
. 3389 / fpsyg . 2019 . 00560
Centelles, L., Assaiante, C., Nazarian, B., Anton, J. L.,
& Schmitz, C. (2011). Recruitment of both the mirror
and the mentalizing networks when observing social
interactions depicted by point-lights: A neuroimaging
study. PLoS One, 6(1), e15749. https://doi . org / 10 . 1371
/ journal . pone . 0015749
Charest, I., Pernet, C., Latinus, M., Crabbe, F., & Belin,
P. (2013). Cerebral processing of voice gender studied
using a continuous carryover FMRI design. Cereb
Cortex, 23(4), 958–966. https://doi . org / 10 . 1093 / cercor
/ bhs090
Cotosck, K. R., Meltzer, J. A., Nucci, M. P., Lukasova,
K., Mansur, L. L., & Amaro, E. (2021). Engagement of
language and domain general networks during word
monitoring in a native and unknown language. Brain Sci,
11(8), 1063. https://doi . org / 10 . 3390 / brainsci11081063
de Gelder, B., de Borst, A. W., & Watson, R. (2015). The
perception of emotion in body expressions. Wiley
Interdiscip Rev Cogn Sci, 6(2), 149–158. https://doi . org
/ 10 . 1002 / wcs . 1335
Deen, B., Koldewyn, K., Kanwisher, N., & Saxe, R. (2015).
Functional organization of social perception and cognition
in the superior temporal sulcus. Cereb Cortex, 25(11),
4596–4609. https://doi . org / 10 . 1093 / cercor / bhv111
Deen, B., Saxe, R., & Kanwisher, N. (2020). Processing
communicative facial and vocal cues in the superior
temporal sulcus. Neuroimage, 221, 117191. https://doi
. org / 10 . 1016 / j . neuroimage . 2020 . 117191
Demenescu, L. R., Mathiak, K. A., & Mathiak, K. (2014).
Age- and gender-related variations of emotion
recognition in pseudowords and faces. Exp Aging Res,
40(2), 187–207. https://doi . org / 10 . 1080 / 0361073X . 2014
. 882210
Ding, X., Gao, Z., & Shen, M. (2017). Two equals one: Two
human actions during social interaction are grouped as
one unit in working memory. Psychol Sci, 28(9), 1311–
1320. https://doi . org / 10 . 1177 / 0956797617707318
Downing, P. E., Jiang, Y., Shuman, M., & Kanwisher, N.
(2001). A cortical area selective for visual processing
of the human body. Science, 293(5539), 2470–2473.
https://doi . org / 10 . 1126 / science . 1063414
Enrici, I., Adenzato, M., Cappa, S., Bara, B. G., &
Tettamanti, M. (2011). Intention processing in
communication: A common brain network for language
and gestures. J Cogn Neurosci, 23(9), 2415–2431.
https://doi . org / 10 . 1162 / jocn . 2010 . 21594
Ethofer, T., Bretscher, J., Gschwind, M., Kreifelts, B.,
Wildgruber, D., & Vuilleumier, P. (2012). Emotional voice
areas: Anatomic location, functional properties, and
structural connections revealed by combined fMRI/DTI.
Cereb Cortex, 22(1), 191–200. https://doi . org / 10 . 1093
/ cercor / bhr113
Ethofer, T., Van De Ville, D., Scherer, K., & Vuilleumier, P.
(2009). Decoding of emotional information in voice-
sensitive cortices. Curr Biol, 19(12), 1028–1033. https://
doi . org / 10 . 1016 / j . cub . 2009 . 04 . 054
Fedorenko, E., Hsieh, P. J., Nieto-Castanon, A., Whitfield-
Gabrieli, S., & Kanwisher, N. (2010). New method
for fMRI investigations of language: Defining ROIs
functionally in individual subjects. J Neurophysiol, 104(2),
1177–1194. https://doi . org / 10 . 1152 / jn . 00032 . 2010
Hasegawa, M., Carpenter, P. A., & Just, M. A. (2002). An
fMRI study of bilingual sentence comprehension and
workload. Neuroimage, 15(3), 647–660. https://doi . org / 10
. 1006 / nimg . 2001 . 1001
Hellbernd, N., & Sammler, D. (2016). Prosody conveys
speaker’s intentions: Acoustic cues for speech act
perception. J Mem Lang, 88, 70–86. https://doi . org / 10
. 1016 / j . jml . 2016 . 01 . 001
Henetz, T. (2017). Don’t hesitate! The length of inter-turn
gaps influences observers’ interactional attributions.
Stanford University. http://purl . stanford . edu / zk615rk5483
Isik, L., Koldewyn, K., Beeler, D., & Kanwisher, N. (2017).
Perceiving social interactions in the posterior superior
temporal sulcus. Proc Natl Acad Sci U S A, 114(43),
E9145–E9152. https://doi . org / 10 . 1073 / pnas . 1714471114
Jacoby, N., Bruneau, E., Koster-Hale, J., & Saxe, R. (2016).
Localizing pain matrix and theory of mind networks
with both verbal and non-verbal stimuli. Neuroimage,
126, 39–48. https://doi . org / 10 . 1016 / j . neuroimage . 2015
. 11 . 025
Jiahui, G., Garrido, L., Liu, R. R., Susilo, T., Barton, J. J. S.,
& Duchaine, B. (2017). Normal voice processing
after posterior superior temporal sulcus lesion.
Neuropsychologia, 105, 215–222. https://doi . org / 10 . 1016
/ j . neuropsychologia . 2017 . 03 . 008
Julian, J. B., Fedorenko, E., Webster, J., & Kanwisher, N.
(2012). An algorithmic method for functionally defining
regions of interest in the ventral visual pathway.
Neuroimage, 60(4), 2357–2364. https://doi . org / 10 . 1016 / j
. neuroimage . 2012 . 02 . 055
Kandylaki, K. D., Nagels, A., Tune, S., Wiese, R.,
Bornkessel-Schlesewsky, I., & Kircher, T. (2015).
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
18
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
Processing of false belief passages during natural
story comprehension: An fMRI study. Hum Brain
Mapp, 36(11), 4231–4246. https://doi . org / 10 . 1002 / hbm
. 22907
Kanwisher, N., & Yovel, G. (2006). The fusiform face area:
A cortical region specialized for the perception of faces.
Philos Trans R Soc Lond B Biol Sci, 361(1476), 2109–
2128. https://doi . org / 10 . 1098 / rstb . 2006 . 1934
Kleiner, M., Brainard, D. H., & Pelli, D. (2007). What’s new in
Psychtoolbox-3? Perception, 36, 1–6. https://doi . org / 10
. 1177 / 03010066070360S101
Kreifelts, B., Ethofer, T., Shiozawa, T., Grodd, W., &
Wildgruber, D. (2009). Cerebral representation of
non-verbal emotional perception: fMRI reveals
audiovisual integration area between voice- and face-
sensitive regions in the superior temporal sulcus.
Neuropsychologia, 47(14), 3059–3066. https://doi . org / 10
. 1016 / j . neuropsychologia . 2009 . 07 . 001
Lahnakoski, J. M., Glerean, E., Slmi, J., Jääskeläinen, L. P.,
Sams, M., Hari, R., & Nummemaa, L. (2012). Naturalistic
fMRI mapping reveals superior temporal sulcus as a hub
for the distributed brain network for social perception.
Front Hum Neurosci, 13(6), 233. https://doi . org / 10 . 3389
/ fnhum . 2012 . 00233
Lakens, D. (2013). Calculating and reporting effect sizes
to facilitate cumulative science: A practical primer for
t-tests and ANOVAs. Front Psychol, 4, 863. https://doi
. org / 10 . 3389 / fpsyg . 2013 . 00863
Landsiedel, J., Daughters, K., Downing, P. E., & Koldewyn,
K. (2022). The role of motion in the neural representation
of social interactions in the posterior temporal cortex.
Neuroimage, 262, 119533. https://doi . org / 10 . 1016 / j
. neuroimage . 2022 . 119533
Latinus, M., McAleer, P., Bestelmeyer, P. E., & Belin, P.
(2013). Norm-based coding of voice identity in human
auditory cortex. Curr Biol, 23(12), 1075–1080. https://doi
. org / 10 . 1016 / j . cub . 2013 . 04 . 055
Lin, N., Wang, X., Xu, Y., Wang, X., Hua, H., Zhao, Y.,
& Li, X. (2018). Fine subdivisions of the semantic
network supporting social and sensory-motor semantic
processing. Cereb Cortex, 28(8), 2699–2710. https://doi
. org / 10 . 1093 / cercor / bhx148
Mar, R. A. (2011). The Neural Bases of Social Cognition and
Story Comprehension. Annual Review of Psychology,
62(1), 103–134. https://doi.org/10.1146/annurev-
psych-120709-145406
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. J Neurosci Methods,
164(1), 177–190. https://doi . org / 10 . 1016 / j . jneumeth . 2007
. 03 . 024
Masson, H. L., & Isik, L. (2021). Functional selectivity for
social interaction perception in the human superior
temporal sulcus during natural viewing. Neuroimage,
245, 118741. https://doi . org / 10 . 1016 / j . neuroimage . 2021
. 118741
Mazaika, P., Whitfield, S., & Cooper, J. C. (2005). Detection
and Repair of Transient Artifacts in fMRI Data. 11th
Annual Meeting of the Organization for Human Brain
Mapping, Toronto, Canada.
McFarquhar, M., McKie, S., Emsley, R., Suckling, J., Elliott,
R., & Williams, S. (2016). Multivariate and repeated
measures (MRM): A new toolbox for dependent and
multimodal group-level neuroimaging data. Neuroimage,
132, 373–389. https://doi . org / 10 . 1016 / j . neuroimage . 2016
. 02 . 053
Mennen, I., Schaeffler, F., & Docherty, G. (2012). Cross-
language differences in fundamental frequency range: A
comparison of English and German. J Acoust Soc Am,
131(3), 2249–2260. https://doi . org / 10 . 1121 / 1 . 3681950
Mitchell, R. L. C., Elliott, R., Barry, M., Cruttenden, A.,
& Woodruff, P. W. R. (2003). The neural response to
emotional prosody, as revealed by functional magnetic
resonance imaging. Neuropsychologia, 41(10), 1410–
1421. https://doi . org / 10 . 1016 / s0028 - 3932(03)00017 - 4
Oldfield R. C. (1971). The assessment and analysis of
handedness: the Edinburgh inventory. Neuropsychologia,
9(1), 97–113. https://doi.org/10.1016/0028-
3932(71)90067-4
Olson, H., Chen, E., Lydic, K., & Saxe, R. (2023). Left-
hemisphere cortical language regions respond equally
to dialogue and monologue. bioRxiv. https://doi . org / 10
. 1101 / 2023 . 01 . 30 . 526344
Oosterhof, N. N., Connolly, A. C., & Haxby, J. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis
of neuroimaging data in Matlab/GNU Octave. Front
Neuroinform, 10, 27. https://doi . org / 10 . 3389 / fninf . 2016
. 00027
Papeo, L. (2020). Twos in human visual perception. Cortex,
132, 473–478. https://doi . org / 10 . 1016 / j . cortex . 2020 . 06
. 005
Papeo, L., & Abassi, E. (2019). Seeing social events:
The visual specialization for dyadic human-human
interactions. J Exp Psychol Hum Percept Perform, 45(7),
877–888. https://doi . org / 10 . 1037 / xhp0000646
Papeo, L., Goupil, N., & Soto-Faraco, S. (2019).
Visual search for people among people. Psychol
Sci, 30(10), 1483–1496. https://doi . org / 10 . 1177
/ 0956797619867295
Papeo, L., Stein, T., & Soto-Faraco, S. (2017). The two-
body inversion effect. Psychol Sci, 28(3), 369–379.
https://doi . org / 10 . 1177 / 0956797616685769
Pernet, C. R., McAleer, P., Latinus, M., Gorgolewski, K. J.,
Charest, I., Bestelmeyer, P. E., Watson, R. H., Fleming, D.,
Crabbe, F., Valdes-Sosa, M., & Belin, P. (2015). The human
voice areas: Spatial organization and inter-individual
variability in temporal and extra-temporal cortices.
Neuroimage, 119, 164–174. https://doi . org / 10 . 1016 / j
. neuroimage . 2015 . 06 . 050
Pijper, J. R. d., & Sanderman, A. A. (1994). On the
perceptual strength of prosodic boundaries and its
relation to suprasegmental cues. J Acoust Soc Am,
96(4), 2037–2047. https://doi . org / 10 . 1121 / 1 . 410145
Ponsot, E., Burred, J. J., Belin, P., & Aucouturier, J. J.
(2018). Cracking the social code of speech prosody
using reverse correlation. Proc Natl Acad Sci U S A,
115(15), 3972–3977. https://doi . org / 10 . 1073 / pnas
. 1716090115
Price, C. J. (2012). A review and synthesis of the first 20
years of PET and fMRI studies of heard speech, spoken
language and reading. Neuroimage, 62(2), 816–847.
https://doi . org / 10 . 1016 / j . neuroimage . 2012 . 04 . 062
Roberts, F., & Francis, A. L. (2013). Identifying a temporal
threshold of tolerance for silent gaps after requests.
J Acoust Soc Am, 133(6), EL471–EL477. https://doi . org
/ 10 . 1121 / 1 . 4802900
Roberts, F., Francis, A. L., & Morgan, M. (2006). The
interaction of inter-turn silence with prosodic cues in
listener perceptions of “trouble” in conversation. Speech
Commun, 48(9), 1079–1093. https://doi . org / https://doi
. org / 10 . 1016 / j . specom . 2006 . 02 . 001
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
19
J. Landsiedel and K. Koldewyn
Imaging Neuroscience, Volume 1, 2023
Robins, D. L., Hunyadi, E., & Schultz, R. T. (2009). Superior
temporal activation in response to dynamic audio-visual
emotional cues. Brain Cogn, 69(2), 269–278. https://doi
. org / 10 . 1016 / j . bandc . 2008 . 08 . 007
Saarela, M. V., & Hari, R. (2008). Listening to humans
walking together activates the social brain circuitry.
Soc Neurosci, 3(3–4), 401–409. https://doi . org / 10 . 1080
/ 17470910801897633
Saxe, R. R., Whitfield-Gabrieli, S., Scholz, J., & Pelphrey,
K. A. (2009). Brain regions for perceiving and reasoning
about other people in school-aged children. Child Dev,
80(4), 1197–1209. https://doi . org / 10 . 1111 / j . 1467 - 8624
. 2009 . 01325 . x
Neuroimage, 30(4), 1414–1432. https://doi . org / 10 . 1016 / j
. neuroimage . 2005 . 11 . 002
Vigneau, M., Beaucousin, V., Herve, P. Y., Jobard, G.,
Petit, L., Crivello, F., Mellet, E., Zago, L., Mazoyer, B., &
Tzourio-Mazoyer, N. (2011). What is right-hemisphere
contribution to phonological, lexico-semantic, and
sentence processing? Insights from a meta-analysis.
Neuroimage, 54(1), 577–593. https://doi . org / 10 . 1016 / j
. neuroimage . 2010 . 07 . 036
Walbrin, J., Downing, P., & Koldewyn, K. (2018). Neural
responses to visually observed social interactions.
Neuropsychologia, 112, 31–39. https://doi . org / 10 . 1016 / j
. neuropsychologia . 2018 . 02 . 023
Schirmer, A., & Adolphs, R. (2017). Emotion perception
Walbrin, J., & Koldewyn, K. (2019). Dyadic interaction
from face, voice, and touch: Comparisons and
convergence. Trends Cogn Sci, 21(3), 216–228. https://
doi . org / 10 . 1016 / j . tics . 2017 . 01 . 001
Scott, S. K., Blank, C. C., Rosen, S., & Wise, R. J. (2000).
Identification of a pathway for intelligible speech in the
left temporal lobe. Brain, 123 Pt 12, 2400–2406. https://
doi . org / 10 . 1093 / brain / 123 . 12 . 2400
Scott, S. K., Rosen, S., Beaman, C. P., Davis, J. P., & Wise,
R. J. (2009). The neural processing of masked speech:
Evidence for different mechanisms in the left and right
temporal lobes. J Acoust Soc Am, 125(3), 1737–1743.
https://doi . org / 10 . 1121 / 1 . 3050255
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster
enhancement: Addressing problems of smoothing,
threshold dependence and localisation in cluster
inference. Neuroimage, 44(1), 83–98. https://doi . org / 10
. 1016 / j . neuroimage . 2008 . 03 . 061
Stevenage, S. V., Hugill, A., & Lewis, H. G. (2012).
Integrating voice recognition into models of person
perception. J Cognit Psychol, 24, 409–419. https://doi
. org / 10 . 1080 / 20445911 . 2011 . 642859
Todorov, A., Olivola, C. Y., Dotsch, R., & Mende-Siedlecki,
P. (2015). Social attributions from faces: Determinants,
consequences, accuracy, and functional significance.
Annu Rev Psychol, 66, 519–545. https://doi . org / 10 . 1146
/ annurev - psych - 113011 - 143831
Van Overwalle, F., & Baetens, K. (2009). Understanding
others’ actions and goals by mirror and mentalizing
systems: A meta-analysis. Neuroimage, 48(3), 564–584.
https://doi . org / 10 . 1016 / j . neuroimage . 2009 . 06 . 009
Vestner, T., Tipper, S. P., Hartley, T., Over, H., &
Rueschemeyer, S. A. (2019). Bound together: Social
binding leads to faster processing, spatial distortion,
and enhanced memory of interacting partners. J Exp
Psychol Gen, 148(7), 1251–1268. https://doi . org / 10 . 1037
/ xge0000545
processing in the posterior temporal cortex. Neuroimage,
198, 296–302. https://doi . org / 10 . 1016 / j . neuroimage . 2019
. 05 . 027
Walbrin, J., Mihai, I., Landsiedel, J., & Koldewyn, K. (2020).
Developmental changes in visual responses to social
interactions. Dev Cogn Neurosci, 42, 100774. https://doi
. org / 10 . 1016 / j . dcn . 2020 . 100774
Walenski, M., Europa, E., Caplan, D., & Thompson, C. K.
(2019). Neural networks for sentence comprehension
and production: An ALE‐based meta‐analysis of
neuroimaging studies. Human Brain Mapping, 40(8),
2275–2304. https://doi.org/10.1002/hbm.24523
Watson, R., Latinus, M., Charest, I., Crabbe, F., & Belin, P.
(2014). People-selectivity, audiovisual integration and
heteromodality in the superior temporal sulcus. Cortex,
50, 125–136. https://doi . org / 10 . 1016 / j . cortex . 2013 . 07 . 011
Wiethoff, S., Wildgruber, D., Kreifelts, B., Becker, H.,
Herbert, C., Grodd, W., & Ethofer, T. (2008). Cerebral
processing of emotional prosody—influence of acoustic
parameters and arousal. Neuroimage, 39(2), 885–893.
https://doi . org / 10 . 1016 / j . neuroimage . 2007 . 09 . 028
Wright, T. M., Pelphrey, K. A., Allison, T., McKeown, M. J.,
& McCarthy, G. (2003). Polysensory interactions along
lateral temporal regions evoked by audiovisual speech.
Cereb Cortex, 13(10), 1034–1043. https://doi . org / 10
. 1093 / cercor / 13 . 10 . 1034
Yang, D. Y.-J., Rosenblau, G., Keifer, C., & Pelphrey, K. A.
(2015). An integrative neural model of social perception,
action observation, and theory of mind. Neurosci
Biobehav Rev, 51, 263–275. https://doi . org / 10 . 1016 / j
. neubiorev . 2015 . 01 . 020
Yang, J. (2014). The role of the right hemisphere in
metaphor comprehension: A meta-analysis of functional
magnetic resonance imaging studies. Human Brain
Mapping, 35(1), 107–122. https://doi.org/10.1002/
hbm.22160
Vigneau, M., Beaucousin, V., Herve, P. Y., Duffau, H.,
Zhang, G., Xu, Y., Zhang, M., Wang, S., & Lin, N. (2021).
Crivello, F., Houde, O., Mazoyer, B., & Tzourio-Mazoyer,
N. (2006). Meta-analyzing left hemisphere language
areas: Phonology, semantics, and sentence processing.
The brain network in support of social semantic
accumulation. Soc Cogn Affect Neurosci, 16(4),
393–405. https://doi . org / 10 . 1093 / scan / nsab003
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00003/2154694/imag_a_00003.pdf by guest on 07 September 2023
20